


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ЛИСКОВАЯ
Алена Анатольевна
Фаунистическое и экологическое разнообразие
панцирных клещей (Acariformes: Oribatei)
в экосистемах Кольского Севера
03.02.04 – зоология
03.02.08 – экология
Автореферат диссертации на соискание учёной степени кандидата
биологических наук
Петрозаводск – 2011
Работа выполнена в Институте проблем промышленной экологии Севера Кольского научного центра Российской академии наук
Научные руководители | доктор биологических наук профессор заслуженный эколог России |
кандидат биологических наук доцент |
Официальные оппоненты доктор биологических наук
профессор
кандидат биологических наук
Ведущая организация Институт проблем экологии и эволюции
им. РАН
Защита диссертации состоится 23 марта 2011 г. в 14 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете Республика Карелия, Петрозаводск, пр. Ленина, 33; эколого-биологический факультет.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета. Электронная версия автореферата размещена на официальном сайте http://www.petrsu.ru.
Автореферат разослан « 15 » февраля 2011 г.
Ученый секретарь диссертационного совета канд. биол. наук |
|
|
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Панцирные клещи или орибатиды (Acariformes: Oribatida) – эволюционно древняя, широко распространенная и чрезвычайно разнообразия в таксономическом и экологическом отношении группа клещей. В почвах северных широт эти микроартроподы преобладают по обилию и видовому разнообразию над представителями мезофауны. Зоо-микробиальные взаимосвязи орибатид с мицелиальными грибами, определяют значимую роль этих клещей в процессах биотрансформации органического вещества в экосистемах полярных и приполярных регионов, включая зональные почвы Кольского Севера, в которых грибы являются ведущей группой микроорганизмов-деструкторов (Евдокимова, Мозгова, 2001).
На территории области, несмотря на небольшую протяженность с юга на север (~400 км), выделяют пять климатических районов (Яковлев, 1961). Здесь выражена зональная смена северотаежного, лесотундрового и тундрового типа растительности. В горах широтной зональности соответствует высотная поясность: горно-таежный пояс сменяется субальпийским (горно-лесотундровым), альпийским (горно-тундровым) и поясом арктических пустынь. В пределах региона на породах разного генезиса сформированы подзолы (иллювиально-железистые, иллювиально-гумусовые, торфяно-подзолы, глееподзолы), подбуры, торфяные почвы, литоземы, криоземы (Переверзев, 2007). В результате развитой горнодобывающей и перерабатывающей промышленности почвы региона подвергаются аэротехногенному загрязнению поллютантами различной природы; на обширных территориях складированы отходы обогащения полезных ископаемых.
Указанные факторы не могут не отражаться на фаунистическом и экологическом разнообразии панцирных клещей, сведения о котором ограничены данными лишь по двум климатическим районам: Терскому берегу и островам Белого моря, насчитывающим немногим более 100 видов (Бызова и др., 1986; Каталог биоты …, 2008) и побережью Баренцева моря, для которого ранее указывалось 79 видов орибатид (Криволуцкий, 1966; Панцирные клещи, 1995; Криволуцкий и др., 1999). Исследования последних лет, направленные на оценку вклада мигрирующих птиц в распространение клещей путем форезии, пополнили список орибатид Кольской тундры 44 видами, населяющими специфические местообитания баренцевоморского побережья – орнитогенные субстраты и гнезда птиц (Лебедева, Криволуцкий, 2003; Лебедева и др., 2006; Лебедева, Лебедев, 2007; Лебедев, 2009). Фауна орибатид основной территории региона, включающей широкий спектр природных и техногенно трансформированных биогеоценозов, а также горные экосистемы, остается практически неизученной.

Цель работы состояла в обобщении разрозненных литературных данных и результатов собственных исследований о фаунистическом и экологическом разнообразии, биоиндикационной значимости панцирных клещей в природных и техногенно трансформированных почвах и техногенных субстратах Мурманской области, а также составлении сводного списка орибатид этого заполярного региона.
Для достижения поставленной цели решали следующие задачи:
1. Дать сравнительную характеристику сообществ орибатид в природных почвах Мурманской области с учетом факторов широтной зональности (почвы тундры, лесотундры и подзоны северной тайги) и высотной поясности (почвы Хибинского горного массива).
2. Сравнить сообщества орибатид в техногенно нарушенных почвах в зонах воздействия промышленных предприятий разного типа (медно-никелевый комбинат, алюминиевый завод). Выявить виды-индикаторы промышленного загрязнения.
3. Исследовать комплексы орибатид в техногенных субстратах на примере самозарастающих и рекультивированных отходах обогащения апатит-нефелиновых руд разного срока складирования.
Научная новизна. Впервые обобщены литературные данные и результаты собственных исследований фауны панцирных клещей в 11 районах Мурманской области, охватывающих естественные (равнинные и горные) и техногенно трансформированные экосистемы тундровой зоны и северотаежной подзоны. Региональный список орибатид увеличен на 79 видов (или 30%), из них 45 видов из 30 родов и 22 семейств указаны впервые. Значительно расширен видовой состав семейств Suctobelbidae, Carabodidae, Oppiidae, наиболее богатых по числу видов в данной области. Установлено влияние широтной зональности и разных механизмов расселения орибатид на их разнообразие, соотношение высших (Brachypylina) и низших (Macropylina) таксонов и соотношение одно-, мало - и многовидовых семейств. Определены виды с широким и локальным распространением в пределах области, а также виды, доминирующие по численности.
Теоретическая значимость. Выявленные виды панцирных клещей дополняют сведения по фауне Мурманской области и значительно расширяют имеющиеся сведения по разнообразию этой группы на территории Европейского Севера России, что важно с точки зрения инвентаризации и сохранения биоразнообразия почв северных регионов. Полученные результаты дают представление о механизмах расселения и формирования фауны клещей в высоких широтах, а также об особенностях экологии орибатид в зональных и техногенно нарушенных почвах Севера.
Практическая значимость. Установлены различные изменения структуры сообществ панцирных клещей в зависимости от степени техногенной нагрузки. Выявлены виды – положительные и отрицательные индикаторы загрязнения почв выбросами алюминиевого и медно-никелевого предприятий, а также виды, устойчивые к воздействию промышленных источников разного типа. Установлена значимость панцирных клещей как индикаторов микологической активности и накопления органического вещества в техногенных субстратах (нефелинсодержащих песках) в процессе первичного почвообразования. Выявлена индикаторная роль орибатид при оценке эффективности способов восстановления территории техногенной пустоши в окрестностях медно-никелевого комбината. Уточненный и дополненный список панцирных клещей Мурманской области опубликован в тематическом выпуске «Зеленый пояс Фенноскандии» Трудов Карельского НЦ РАН.
Апробация работы. Основные результаты исследований были доложены и получили положительную оценку на 8 научных конференциях с международным участием: «Наука и образование» (Мурманск 2008); «Актуальные проблемы сохранения биоразнообразия в экстремальных условиях северного климата» (Кировск 2008); «Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт» (Мурманск 2009); «Сбалансированное природопользование» (Апатиты 2009); «Биогеграфия почв» (Москва 2009); «Современные проблемы загрязнения почв» (Москва 2010); «Экологические проблемы северных регионов и пути их решения» (Апатиты 2010); XV Всероссийское совещание по почвенной зоологии (Москва 2008).
Публикации. По результатам диссертации опубликовано 13 работ, в том числе две статьи в рецензируемых журналах из списка, рекомендованного ВАК.
Структура и объем диссертации. Диссертация состоит из введения, 7 глав, выводов, 6 приложений с видовым списком панцирных клещей Мурманской области, перечня цитируемой литературы, включающего 233 источника, в т. ч. 92 на иностранных языках. Работа изложена на 178 страницах, содержит 57 рисунков, 55 таблиц.
Благодарности. Автор выражает искреннюю благодарность д. б.н., профессору и к. б.н., доценту за руководство работой, идеи и постоянную помощь при подготовке диссертации, за предоставленные данные по микробиологическим и физико-химическим свойствам исследованных почв; к. б.н. за предоставленные данные и консультации.
Особую благодарность за таксономическое определение панцирных клещей, предоставленные данные и персональные консультации выражаем к. б.н. , к. г.н. , к. г.н. и .
СОДЕРЖАНИЕ РАБОТЫ
1. СИСТЕМТИческое положение И ЭКОЛОГИЯ ПАНЦИРНЫХ клещей
Приведены сведения о систематическом положении, таксономическом разнообразии, особенностям биологи и экологии панцирных клещей, их использовании в зоологической индикации почв. Показана степень изученности фауны панцирных клещей на территории Мурманской области.
2. ХАРАКТЕРИСТИКА МУРМАНСКОЙ ОБЛАСТИ
Рассмотрено разнообразие природных условий и охарактеризованы типы промышленного загрязнения почв Мурманской области как факторов, определяющих биоразнообразие региональной фауны панцирных клещей.
3. ОБЪЕКТЫ И МЕТОДЫ
Исследования охватили 11 районов Мурманской области от побережья Баренцева моря (тундрово-лесотундровая флористическая область, районы 1-3) до побережья Кандалакшского залива Белого моря (районы 10, 11), включая природные, техногенные и горные экосистемы центральной части региона (подзона северной тайги, районы 4-9) (рис. 1, табл. 1). Собственные исследования выполнены в 4 районах: 4-7. Материалы по районам 1, 2, 8, 9 предоставлены сотрудниками ИППЭС Кольского НЦ РАН. По районам 3, 10, 11 использованы литературные данные (Криволуцкий, 1966; Бызова и др., 1986; Панцирные клещи, 1995; Криволуцкий и др., 1999; Лебедева, Криволуцкий, 2003; Лебедева и др., 2006; Лебедева, Лебедев, 2007; Лебедев, 2009). Видовой состав панцирных клещей приведен для 9 районов: 1, 2, 3, 5, 6, 8, 9, 10, 11. Виды, выявленные в районах 4 и 7, будут идентифицированы позднее. Всего исследовано 43 биотопа.
| Рис. 1. Районы исследований: 1 –пос. Видяево; 2 – пос. Териберка; 3 – пос. Дальние Зеленцы; 4, 5 –участки в 1 и 5 км от медно-никелевого комбината «Североникель» (г. Мончегорск); 6 – Хибинский горный массив, 7 –хранилища нефелинсодержащих песков, 8 – пос. Уполокша, зональные сосняки, 9 – зона в 2 км от алюминиевого завода (КАЗ, г. Кандалакша), 10 – беломорские о-ва Кандалакшского заповедника, 11 – Терский берег Белого моря. |
Таблица 1. Характеристика исследованных районов
№ | Коорд. | Экосистемы, почвы |
Тундрово-лесотундровая флористическая область | ||
1 | 69°19′ с. ш. 32°52′ в. д. | Березняк лесотундровый кустарничково-лишайниковый; вершина скалы, 60 м над ур. м. Почвы: Al-Fe-гумусовые подзолы, торфяно-подзолы. |
2 | 69°10' с. ш. 35°08' в. д. | Горные тундра лишайниково-кустарничковая и березняк лесотундровый воронично-лишайниковый, 40-60 м над ур. м. Подзолы Al-Fe-гумусовые. |
3 | 69°07' с. ш. 36°03' в. д. | Тундра кустарничково-лишайниковая, 50-80 м над ур. м. Подзолы Al-Fe-гумусовые оторфованные. |
Подзона северной тайги | ||
4 | 67°56' с. ш. 32°49' в. д. | Площадки по восстановлению техногенной пустоши в 1 км от комбината «Североникель», 138-177 м над ур. м. |
5 | 67°34' с. ш. 34°17' в. д. | Импактная зона в 5 км от комбината «Североникель», 80 м над ур. м. Подзолы Al-Fe-гумусовые оторфованные, сильно эродированные. В верхнем слое содержится (мг/кг): Ni , Cu , Co 300-350, сульфат-ионов 400-600. Зольность достигает 80-90%, pH = 3.5. |
6 | Хибинский горный массив (ХГМ), 6 биотопов | |
67°38' с. ш. 33°39' в. д. | Гора Вудъяврчорр, склон северо-восточной экспозиции. Субальпийский пояс березового криволесья (В-СВЭ-бк), 393 м над ур. м. Подзол иллювиально-многогумусовый. | |
67°38' с. ш. 33°39' в. д. | Гора Вудъяврчорр, склон северо-восточной экспозиции. Горная тундра кустарничковая, 434 над ур. м. (В-СВЭ-т/куст). Почва скрытоподзолистая иллювиально-многогумусовая оторфованная. | |
67°38' с. ш. 33°40' в. д. | Гора Вудъяврчорр, южный склон. Предгорная ерниковая тундра (В-ЮЭ-т/ерн), 324 м над ур. м. | |
67°10' с. ш. 33°12' в. д. | Западная часть ХГМ, правый берег реки Малая Белая, горно-таежный пояс (ХГМ-ЗЧ-гт). | |
67°14' с. ш. 33°34' в. д | Восточная часть ХГМ, пойма реки Тульйок, горно-таежный пояс (ХГМ-ВЧ-гт), 280 м над ур. м. | |
67°10' с. ш. 38°00' в. д. | Гора Ловчорр, западный склон. Тундра каменистая с фрагментарным лишайниково-кустарничковым покровом (Л-ЗЭ-т/кам). | |
7 | 67°35' с. ш. 33°35' в. д. | Хранилища с нефелинсодержащими песками разного срока хранения: свеженамытые (срок экспозиции <1 года); самозарастающие мхами и закрепленные методом посева многолетних злаков более 10-, 20- и 30-ти лет назад; рекультивированные более 40 лет назад – новообразованные почвы под лесной и луговой растительностью. |
8 | 67°33' с. ш. 31°54' в. д. | Зональный сосняк вороничный. Подзолы Al-Fe-гумусовые на песчаной морене. Содержание углерода – 52% от сухого вещества, азота – 1.43%. |
9 | 67°09' с. ш. 32°24' в. д. | Сосняк вороничный в зоне сильного загрязнения в 2 км от КАЗ, функционирующего с 1951 г. Древостой разреженный, с отмершими ветками и пожелтевшей хвоей в нижней и средней частях кроны. Подзолы Al-Fe-гумусовые в состоянии техногенной трансформации. Содержание фтора в подстилке до 1200 мг/кг. Органическое вещество подстилки обеднено углеродом и азотом: содержание С-27% от сухого вещества, N-0.63%. Зольность повышена до 60%, величина pH – до 5.5. |
10, 11 | 66o34' с. ш. 33o08' в. д. | О-ва Северного Архипелага и Терское побережье Белого моря. Сосновые, еловые, березовые леса, приморские луга, полоса галофитов, наскальная растительность (31 биотоп). Подзолы, торфяно-подзолы. |
Клещи были извлечены из почвенных проб размером 25´25 см, отобранных из органогенного горизонта 0-5 см, а также из образцов с растительными остатками (лесная подстилка, листья березы, хвоя сосны), заложенными в органогенный горизонт в мешочках из нейлоновой сетки с диаметром отверстий 1×1 мм. В районах 4 и 6 образцы отбирали металлическим цилиндром диаметром 10 см и высотой 5 см. Для выгонки клещей использовали электропрогрев по методу эклекторов Тулльгрена в течение нескольких суток до полного высушивания субстратов. В период с мая 2007 по июнь 2010 года выполнено 16 полевых выездов. Собрано и обработано более 860 образцов. Экстрагировано и определено около 19.4 тыс. экземпляров панцирных клещей.
Одновременно был выполнен анализ физико-химических свойств субстратов: температуры, влажности, зольности, кислотности (~4 тыс. определений). Связь фаунистических и микробиологических показателей, почвенных свойств и показателей ферментативной активности проанализированы по результатам корреляций. Сходство видового состава клещей в исследованных районах оценивали с помощью кластерного анализа. В качестве базовой меры сходства взят индекс общности Серенсена Is. Рассчитывали прогнозное разнообразие орибатид по формуле: Ds= S×(2-0.08n), n ≥ 11, где n – число исследованных районов, S – число видов в данном районе (Криволуцкий и др., 1999). Данные обрабатывали в программах Microsoft Excel (описательная статистика, корреляционный анализ) и Statistica 7.0 (кластерный анализ). Использовали трофическую классификацию панцирных клещей (Shuester, 1956; Wallwork, 1958; Xavier, Haq, 2007) и их подразделение на жизненные формы (Криволуцкий и др., 1990). Типы ареалов приведены в соответствии с системой Субиаса (Subias, 2010).
4. ФАУНА ПАНЦИРНЫХ КЛЕЩЕЙ ЗОНАЛЬНЫХ ПОДЗОЛОВ КОЛЬСКОГО СЕВЕРА
4.1. Панцирные клещи в почвах зональной тундры. Список панцирных клещей в окрестностях пос. Дальние Зеленцы (район 3), обобщенный по разным источникам, насчитывает 123 вида из 72 родов, 38 сем. В результате исследований районов 1 и 2 северо-западнее района 3 на побережье Баренцева моря, состав орибатид Кольской тундры пополнился 7 новыми видами и родом Diapterobates. Теперь он насчитывает 143 вида, 78 родов, 38 сем. Наиболее разнообразны 17 семейств (или 45%) с числом видов от 4 до 10, в первую очередь – сем. Brachychthoniidae. Треть семейств одновидовые.
В районе 1 в лесотундровых подзолах под березняками найдено 14 видов орибатид, принадлежащих к 12 родам, 11 сем. Виды Ph. anonymus, H. gibba и C. minusculus указываются для Мурманской области впервые. За исключением сем. Carabodidae (3 вида рода Carabodes) все семейства включали по 1 роду с единственным видом, что соответствует организации сообществ педобионтов, находящихся за пределами экологического оптимума (Ганин, 2006, 2009). Экстремальность условий местообитания подтверждалась супердоминированием наиболее эврибионтного среди орибатид вида T. velatus – 65% (рис. 2).
| Рис. 2. Структура доминирования панцирных клещей в почвах зональной тундры в окрестностях поселка Видяево. |

В горно-тундровых и лесотундровых формациях района 2 обнаружено 27 видов из 24 родов, 18 сем. При доминировании вида T. velatus (28%) шире был спектр как доминантов (8 видов), так и видов c обилием менее 1% каждый (19 видов). Структура сообщества была полидоминантной (рис. 3). Четыре малочисленных вида – Ph. crenophilus, M. translamellata, E. acromios и D. humeralis выявлены в регионе впервые. Таксономический статус нескольких интересных экземпляров требует уточнения. Возможно, в эти районы с Нордкапским течением проникают виды, характерные для атлантического побережья Европы.
| Рис. 3. Структура доминирования панцирных клещей в почвах зональной тундры в окрестностях поселка Териберка. |
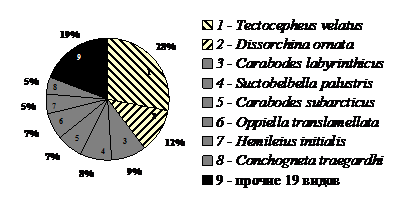
Общими для районов 1 и 2 были 6 видов (Is = 29%): T. velatus, N. coronata, C. traegardhi, H. initialis, C. subarcticus, C. forsslundi. Лесной ксерофильный вид C. forsslundi, известный из оперения птиц (Лебедева, 2005), отмечен только в этих районах (табл. 2).
Для трех тундровых районов побережья Баренцева моря общими оказались лишь 2 вида: T. velatus, широко распространенный по территории Мурманской области, и C. subarcticus, тяготеющий к зональным и горным тундрам региона и не выявленный в широком спектре островных местообитаний Белого моря. Подавляющее большинство – 125 видов (или 88%) встречены лишь в одном из трех районов, для трети из них (40 видов или 32%) известна форезия на птицах. Потенциально птицами может переноситься 55 видов или ~40% орибатид Кольской тундровой зоны.
Таблица 2. Панцирные клещи, впервые указанные для Мурманской области
Семейство | Род | Вид | Район |
Achipteriidae | Achipteria | A. coleoptrata (Linneus 1758) | 8, 9 |
Parhypochthoniidae | Parhypochthonius | P. aphidinus (Berlese 1904) | 9 |
Camisiidae | Camisia | C. borealis (Hermann 1804) | 5 |
Heminothrus | H. septentrionalis (Sellnick 1944) | 6 | |
Euphthiracaridae | Mesotritia | M. flagelliformis (Ewing 1909) | 9 |
Euphthiracarus | E. monodactylus (Willmann 1919) | 8 | |
Microtritia | M. minima (Berlese 1904) | 1 | |
Trhypochthoniidae | Trhypochthonius | T. cladonicola (Willmann 1919) | 9 |
Oppiidae | Medioppia | M. globosa (Mihelčič 1956) | 9 |
Moritzoppia | M. unicarinata (Paoli 1908) | 8, 9 | |
Scheloribatidae | Paraleius | P. leontonychus (Berlese 1910) | 9 |
Hemileius | Hemileius sp. | 6 | |
Phenopelopidae | Eupelops | E. acromios (Hermann 1804) | 2 |
E. occultus (C. L. Koch 1836) | 5, 8, 9 | ||
Haplozetidae | Protoribates | P. capucinus (Berlese 1908) | 6 |
Phthiracaridae | Phthiracarus | Ph. anonymus (Grandjean 1933) | 1 |
Ph. crenophilus (Willmann 1951) | 2 | ||
Ph. laevigatus (C. L. Koch 1844) | 2, 8 | ||
Ph. piger (Scopoli 1763) | 8 | ||
Hermanniidae | Hermannia | H. gibba (C.L.Koch 1840) | 1 |
Carabodidae | Carabodes | C. coriaceus (C.L. Koch 1835) | 8 |
C. forsslundi (Sellnick 1953) | 1, 2 | ||
C. minusculus (Berlese 1923) | 1 | ||
C. reticulatus (Berlese 1913) | 8 | ||
Liacaridae | Liacarus | Liacarus sp. | 2 |
Suctobelbidae | Suctobelbella | S. alloenasuta (Moritz 1971) | 8 |
S. baloghi (Forsslund 1958) | 8 | ||
S. falcata (Forsslund 1941) | 8, 9 | ||
S. forsslundi (Strenzke 1950) | 8, 9 | ||
S. latirostris (Strenzke, 1950) | 1, 9 | ||
S. perforata (Strenzke 1950) | 9 | ||
S. singularis (Strenzke 1950) | 8, 9 | ||
S. subtrigona (Oudemans 1900) | 8, 9 | ||
Peloppiidae | Ceratoppia | C. quadridentata (Haller 1882) | 8 |
Ceratozetidae | Ceratozetella | Ceratozetella sp. | 6 |
Diapterobates | D. humeralis (Hermann 1804) | 2 | |
Diapterobates sp. | 6 | ||
Chamobatidae | Chamobates | Chamobates sp. | 6 |
Damaeidae | Belba | Belba sp. | 6, 8, 9 |
Epidamaeus | Epidamaeus sp. | 6, 8, 9 | |
Eremaeidae | Euеremaeus | E. silvestris (Forsslund 1957) | 6, 8 |
Galumnidae | Pergalumna | P. dorsalis (C. L. Koch 1841) | 8, 9 |
Galumna | Galumna sp. | 6 | |
Nanhermanniidae | Nanhermannia | N. sellnicki (Forsslund 1958) | 6 |
Tectocepheidae | Tectocepheus | T. velatus sarekensis (Trägårdh 1910) | 6 |
22 семейства | 30 родов | 45 видов | 6 |
Примечание. Жирным шрифтом выделены впервые указанные семейства и роды.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
