

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Однако при таком выводе крайне важно различать параметры разнообразия на уровне, говоря языком биологов, высших и низших таксонов. Можно, в частности, выделить социальные «таксоны» более высокого уровня (на уровне семейств, отрядов или даже классов и типов в биологии), такие, например, как племена, вождества, государства и т. п., представленные в каком-то спектре вариаций-видов[26]. Но по мере развития более ароморфных таксонов высокого уровня (например, при эволюционировании сложных вождеств и их аналогов в государства) в рамках сохраняющегося прежнего таксона количество таких вариаций-видов мог-
ло значительно сократиться. Иными словами, количество высших таксо-
нов до поры до времени в тенденции росло (а количество низших таксонов архаических социальных типов с определенного времени начало в тенденции сокращаться).
В социальной макроэволюции появление нового по уровню ароморфного таксона обычно в тенденции и на больших временных отрезках ведет к тому, что в рамках старого таксона (например, негосударственных форм политий) разнообразие начинает с определенного момента уменьшаться, а в рамках нового таксона (например, государств) это разнообразие, наоборот, растет. Но сами старые таксоны высшего порядка могут оставаться как действующая реальность еще очень долго.
Действительно, скажем, появление простых вождеств (и их аналогов) не привело к исчезновению независимых общин; появление сложных
вождеств (и их аналогов) не привело к исчезновению простых вождеств; появление ранних государств (и их аналогов) не привело к исчезновению вождеств и независимых общин; появление развитых государств (и их аналогов) не привело к исчезновению независимых общин, вождеств, ранних государств и их аналогов. Даже появление в XVIII – начале XIX вв. первых зрелых государств и их аналогов не привело к немедленному исчезновению независимых общин, простых и сложных вождеств, ранних и развитых государств и их аналогов.
В результате максимального уровня разнообразие политических систем (на уровне высших таксонов) достигло в XIX в., когда мы видим одновременное существование всех вышеназванных политических форм. То же самое максимальное разнообразие мы видим в этот период не только по политическим, но и по всем другим параметрам. К середине XIX в. было достигнуто максимальное разнообразие производственно-экономичес-ких форм – одновременное существование самых разных видов неспециализированных бродячих охотников-собирателей, специализированных осед-лых охотников-собирателей, ранних экстенсивных земледельцев, кочевых скотоводов, обществ, специализирующихся на торговле и/или ремесле, развитых интенсивных земледельцев и первых (но уже вполне сложившихся) промышленных обществ. Мы видим максимальное разнообразие и, скажем, религиозных форм (сосуществование самых разных анимистических, тотемистических, фетишистских, шаманистских, политеистических и т. п. религий наряду с самыми разнообразными деноминациями и сектами мировых и синкретических религий) и т. п.
Систематическое же сокращение разнообразия политических, социальных, экономических и т. п. форм/высших таксонов началось только со второй половины XIX в., когда по этому показателю социальная макроэволюция начала радикально отличаться от биологической[27].
Тем не менее, в социальной эволюции процесс роста разнообразия можно увидеть и в последние десятилетия, однако он достигается иными способами, чем в биологической эволюции: дифференциацией институтов, отношений, социальных групп, ростом разнообразия специализаций в рамках одной профессии, в номенклатуре артефактов, информации и т. п. Эту тенденцию можно обозначить как рост разнообразия результатов человеческой деятельности. Нет никакого сомнения, что рост этот весьма значителен, причем темпы его нарастают. В то же время, как уже отмечалось, в социальной макроэволюции последних десятилетий постоянно действует и противоположная мощнейшая тенденция на вытеснение и унификацию (культур, языков, религий, институтов, вкусов и т. п.).
В современную эпоху процесс глобализации приводит к сокращению этнического и культурного разнообразия.
Рост разнообразия и многообразия форм имеет своим непосредственным результатом и повышение возможностей появления новых ароморфозов. -Ресовский, и считают, что такой рост ведет в целом к усложнению всей биосферы и, соответственно, к усложнению отношений со средой каждой группы организмов (Тимофеев-Ресовский и др. 1969: 282). Новые перспективные таксоны раньше или позже приобретали особые, ароморфные характеристики, которые позволяли им использовать эти преимущества в более широком аспекте. Таким образом, хотя такие широкие ароморфные адаптации и были редким явлением, однако их частота могла увеличиваться по мере накопления разнообразия, поскольку в плотной конкурентной среде чаще рождались необычные ответы на обычные вызовы среды. Формирование крупных ароморфозов происходит на фоне гибели и неудачных эволюционных попыток многих организмов (обществ) и групп.
а) б)
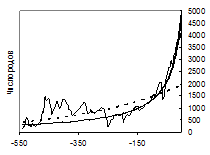
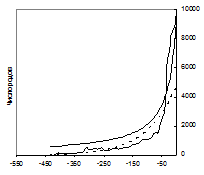
Рис. 1. Рост разнообразия (числа родов) морских (а) и континентальных (б) организмов в течение фанерозоя (542–0 млн лет назад). По горизонтальной оси – время в млн лет назад. Пунктиром показан экспоненциальный тренд, толстой линией – гиперболический тренд (подробнее см.: Марков, Коротаев 2007; 2008; 2009; Гринин, Марков, Коротаев 2008: Приложение 5).
Правило «платы» за ароморфный прогресс
(вместо заключения)
В частности, «как показывает палеонтологическая летопись, из одной адаптивной зоны в другую попадают лишь отдельные сравнительно немногочисленные группы. Этот переход обычно осуществляется с большой (эволюционной) скоростью, причем многие группы гибнут в этих интерзональных промежутках, не достигнув новых оптимальных адаптивных зон. Но зато даже единственная ветвь, попадая в новую адаптивную зону, вступает в новый период аллогенеза» (Тимофеев-Ресовский и др. 1969: 224) (речь идет скорее не об уровне изменений, а об их направлении, то есть об адаптивной радиации, или кладогенезе. – Авт.)».
Если же говорить о социальной эволюции, то это, образно говоря, не широкая лестница, по которой раньше или позже могут подняться в одном направлении и самостоятельно все общества, а сложнейший лабиринт, ароморфный выход из которого без заимствований находят лишь некоторые (и то только на каком-то определенном этапе, а в конечном счете найти «выход из лабиринта» совсем без всяких заимствований не смогло ни одно общество). Иными словами, далеко не каждое конкретное общество является повторением в малом масштабе основной линии арогенного эволюционного развития, а только некоторые из них и лишь на отдельных периодах макроэволюции (и то с большими оговорками). Дело в том, что эволюционный прорыв к качественно новому уровню (ароморфозу) в одном месте (обществе) на протяжении почти всей истории человечества мог состояться только за счет гибели, стагнации, движения вбок и т. п. множества других обществ.
Здесь можно проследить определенное сходство с биологической эволюцией. Стоит вспомнить, например, многочисленные и в конечном счете неудачные «попытки» прокариот перейти к многоклеточности и не менее многочисленные, но отчасти удачные такие же попытки эукариот. В периоды массовых вымираний происходит «предварительный отбор» более жизнеспособных и устойчивых таксонов. Это также означает, что вымершие таксоны освобождают эволюционное пространство для новых потенциальных лидеров, которые в результате получают лучшие стартовые условия, чем таковые были до вымираний.
Таким образом, мы полагаем, что возникновение перспективных морфологических форм, институтов, отношений, с одной стороны, объясняется внутренними свойствами организмов и обществ, но с другой – это всегда результат того, что в наличии имеется достаточно иных форм, эволюционные «удачи» и «неудачи» которых и подготовили, в конце концов, «удачный» вариант.
Библиография
Алексеев А. С. 1998. Массовые вымирания в фанерозое: дис. ... д-ра геол.-минерал. наук. М.: Изд-во МГУ.
2001. Античная философия. М.: Высшая школа.
Бердников В. А. 1990. Основные факторы макроэволюции. Новосибирск: Наука.
1990. Археологический словарь. М.: Прогресс.
Валентайн Дж. 1981. Эволюция многоклеточных растений и животных. В: Майр, Э. и др., Эволюция (перевод темат. вып. журнала Scientific American), c. 149−172. М.: Мир.
1972. Протестантская этика и дух капитализма. М: ИНИОН АН СССР.
Н. 1961. Неравномерность темпов преобразования органов пищевартельной системы грызунов и принцип компенсации функций. Доклады АН СССР 136/6: 1494−1497.
1999. Развитие эволюционных идей в биологии. М.: Прогресс-Традиция.
Георгиевский А. Б. 1974. Проблема преадаптации. Историко-критическое исследование. Л.: Наука.
2000. Ранняя Малая Азия и Хеттское царство. В: Якобсон, В. А. (ред.), История Востока. 1. Восток в древности, с. 113–127. М.: Вост. лит-ра.
2003. Термодинамическая теория структуры, устой-чивости и флуктуации. М.: Едиториал УРСС.
Н. 1977. Ранний железный век. М.: МГУ.
1991. Эволюционный процесс. М.: Мир.
Гринин Л. Е. 1997–2001. Формации и цивилизации. [Книга печаталась в журнале Философия и общество с 1997 по 2001 г.]
Гринин Л. Е. 1997а. Формации и цивилизации. Гл. 1. Философия и общество 1: 10–88.
Гринин Л. Е. 1997б. Формации и цивилизации. Гл. 2. Философия и общество 2: 5–89.
Гринин Л. Е. 1999. Формации и цивилизации. ГлФилософия и общество 2: 5–52.
Гринин Л. Е. 2003а. Производительные силы и исторический процесс. 2-е изд. Волгоград: Учитель.
Гринин Л. Е. 2003б. Философия, социология и теория истории. 3-е изд. Волгоград: Учитель.
Гринин Л. Е. 2007а. Государство и исторический процесс: Политический срез исторического процесса. М.: КомКнига/URSS.
Гринин Л. Е. 2007б. Государство и исторический процесс. Эпоха образования государства: общий контекст социальной эволюции при образовании государства. М: КомКнига/URSS.
Гринин Л. Е. 2007в. Государство и исторический процесс. Эволюция государственности: от раннего государства к зрелому. М: КомКнига/URSS.
Гринин Л. Е. 2007г. Зависимость между размерами общества и эволюционным типом политии. История и Математика: анализ и моделирование социально-исторических процессов / Ред. А. В. Коротаев, С. Ю. Малков, Л. Е. Гринин,
с. 263–303. М.: КомКнига/URSS.
Гринин Л. Е. 2007д. Проблемы анализа движущих сил исторического развития, общественного прогресса и социальной эволюции. Философия истории: проблемы и перспективы / Ред. Ю. И. Семенов, И. А. Гобозов, Л. Е. Гринин, с. 148–247. М.: КомКнига/УРСС.
Гринин Л. Е. 2008. О роли личности в истории. Вестник РАН 78(1): 42–47.
Гринин Л. Е., Коротаев А. В. 2007а. Политическое развитие Мир-Системы: формальный и количественный анализ. История и математика: макроисторическая динамика общества и государства / Ред. С. Ю. Малков, ,
, с. 49–101. М.: КомКнига/URSS.
Гринин Л. Е., Коротаев А. В. 2007б. Социальная макроэволюция и исторический процесс (к постановке проблемы). Философия и общество 2: 19–66; 3: 5–48;
4: 17–50.
Гринин Л. Е., Коротаев А. В. 2008а. История и макроэволюция. Историческая психология и социология истории 2: 59–86
Гринин Л. Е., Коротаев А. В. 2008б. Макроэволюция и Мир-Система: новые грани концептуализации. История и современность 1: 3–31.
Гринин Л. Е., Коротаев А. В. 2009. Социальная макроэволюция. Генезис и трансформации Мир-Системы. М.: Изд-во ЛКИ/URSS.
Гринин Л. Е., Коротаев А. В., Малков С. Ю. 2008. Математические модели социально-демографических циклов и выхода из «мальтузианской ловушки»: некоторые возможные направления дальнейшего развития. Проблемы математической истории: Математическое моделирование исторических процессов / Ред. Г. Г. Малинецкий, А. В. Коротаев, с. 78–117. М.: ЛИБРОКОМ/URSS.
, , 2008. Макроэволюция в живой природе и обществе. М.: УРСС.
1972. Древнейшие цивилизации Мезоамерики. М.: Наука.
Ш. 1972. Учение об эволюционном прогрессе (теория ароморфоза). Тбилиси: Мецниереба.
1991. Происхождение видов путем естественного отбора или охранение благоприятных рас в борьбе за жизнь. СПб.: Наука.
В. 1977. Регион Юго-Восточной Азии с древнейших времен до XV в. Юго-Восточная Азия в мировой истории / Ред. С. Н. Ростовский, Э. О. Берзин, В. Ф. Васильев, А. Р. Вяткин, Г. И. Левинсон, с. 9–69. М.: Наука.
1986. Эволюция организации ДНК приматов. Эволюция генома / Ред. Г. Доувер, Р. Флейвелл, с. 119–138. М.: Мир.
А. 1954. Олигомеризация гомологичных органов как один из главных путей эволюции животных. Л.: Изд-во ЛГУ.
Дьяконов И. М. 2004. Малая Азия, Армянское нагорье и Закавказье в первой половине I тысячелетия до н. э. (Урарту, Фригия, Лидия). История древнего Востока. От ранних государственных образований до древних империй / Ред. А. В. Седов, с. 395–420. М.: Вост. лит-ра.
2005. История Европы. М.: Транзиткнига.
1971. Возникновение государства у фракийцев. М.: Наука.
Иорданский Н. Н. 1977. Неравномерность темпов макроэволюции и ключевые ароморфозы. Природа 6: 3.
Иорданский Н. Н. 2001. Эволюция жизни. М.: Академия.
Кабо В. Р. 1980. У истоков производящей экономики. Ранние земледельцы / Ред. Н. А. Бутинов, А. М. Решетов, с. 59–85. Л.: Наука.
1986. Первобытная доземледельческая община. М.: Наука.
М. 2000. Проблемы, парадоксы и перспективы эволюционизма. Альтернативные пути к цивилизации / Ред. Н. Н. Крадин, А. В. Коротаев, Д. М. Бондаренко, В. А. Лынша, с. 6–23. М.: Логос.
, 1988. Кельты в Европе в первой половине
I тыс. до н. э. История Европы. 1. Древняя Европа / Ред. Е. С. Голубцова,
с. 203–212. М.: Наука.
Коротаев А. В. 1997. Факторы социальной эволюции. М.: Ин-т востоковедения РАН.
Коротаев А. В. 1999. Объективные социологические законы и субъективный фактор. Время мира 1: 204–233.
Коротаев А. В. 2003. Социальная эволюция: факторы, закономерности, тенденции. М.: Вост. лит-ра.
Коротаев А. В., , 2007. Законы истории. Вековые циклы и тысячелетние тренды. Демография. Экономика. Войны. М.: КомКнига/URSS.
Коротаев А. В., Малков А. С., Халтурина Д. А. 2007. Законы истории: Математическое моделирование развития Мир-Системы. Демография, экономика, культура. М.: КомКнига/URSS.
1982. История жизни. Киев: Наукова думка.
Красилов В. А. 1984. Теория эволюции: Необходимость нового синтеза. Эволюционные исследования: Макроэволюция / Ред. , с. 4–12. Владивосток: ДНВЦ АН СССР.
1990. Человек и природа в Китае. М.: Наука.
1974. Популяции, виды и эволюция. М.: Мир.
Майр Э. 1981. Эволюция. В: Майр, Э. и др., Эволюция (перевод темат. вып. журнала Scientific American), с. 11−32. М.: Мир.
1997. Религии человечества. М.; СПб.: Рудомино.
1937. Промышленная революция XVIII столетия в Англии. М.: Соцэкгиз.
Марков А. В., Коротаев А. В. 2007. Динамика разнообразия фанерозойских морских животных соответствует модели гиперболического роста. Журнал общей биологии 68/1: 3–18.
Марков А. В., Коротаев А. В. 2008. Гиперболический рост разнообразия морской и континентальной биот фанерозоя и эволюция сообществ. Журнал общей биологии 69/3: 175–194.
Марков А. В., Коротаев А. В. 2009. Гиперболический рост в живой природе и обществе. М.: УРСС.
Марков А. В., М. 2005. Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы. Палеонтологический журнал 66/4: 3–18.
Марков А. В., 1998. Количественные закономерности макроэволюции. Опыт применения системного подхода к анализу развития надвидовых таксонов. М.: Геос.
1967. Значение воззрений на учение о прогрессе и регрессе в эволюции животных для современной биологии. В: Северцов, А. Н. Главные направления эволюционного прогресса, с. 140–172. М.: Изд-во МГУ.
Мелларт Дж. 1982. Древнейшие цивилизации Ближнего Востока. М.: Наука.
Николис Г., Пригожин И. 1979. Самоорганизация в неравновесных системах. М.: Мир.
Николис Г., Пригожин И. 2003. Познание сложного. Введение. М.: Едиториал УРСС.
В. 1995. Исторические типы общественного воспроизводства: политэкономия мирового исторического процесса. М.: Вост. лит-ра.
Парсонс Т. 2000. О структуре социального действия. М.: АкадемПроект.
1956. К вопросу о роли личности в истории. Избранные философские произведения: в 4 т. Т. 2, с. 300–334. М.: Гос. изд-во полит. лит-ры.
2002. От существующего к возникающему. М.: URSS.
2003. Порядок из хаоса. Новый диалог человека с природой. М.: URSS.
Ратнер В. А. 1992. Блочно-модульный принцип организации и эволюции молекулярно-генетических систем управления (МГСУ). Генетика 28/2: 5–24.
А., А., , Соловьев В. В., 1985. Проблемы теории молекулярной эволюции. Новосибирск: Наука.
Раутиан А. С. 1988а. Палеонтология как источник сведений о закономерностях и факторах эволюции. Современная палеонтология 2: 76–118.
Раутиан А. С. 1988б. Словарь терминов и наименований. Современная палеонтология 2: 356–372. М.: Недра.
Раутиан А. С. 2006. Букет законов эволюции. Эволюция биосферы и биоразнообразия. К 70-летию А. Ю. Розанова / Ред. , с. 20–38. М.: КМК.
2003. Ископаемые бактерии, седиментогенез и ранние стадии эволюции биосферы. Палеонтологический журнал 6: 41−49.
Северцов A. Н. 1935. Модусы филэмбриогенеза. Зоологический журнал 14/1: 1−8.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
