

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 631.847:631.522
© 2005
, кандидат химических наук,
, кандидат биологических наук,
, кандидат сельскохозяйственных наук,
, аспирант, , магистры,
Полтавская государственная аграрная академия,
профессори, доктор,
Аграрный Университет BOKU, Вена, Австрия
БИОЛОГИЧЕСКАЯ ФИКСАЦИЯ АЗОТА РАЗЛИЧНЫМИ СОРТАМИ ЛЮЦЕРНЫ
В УСЛОВИЯХ ВОДНОГО СТРЕССА. ТЕПЛИЧНЫЙ ЭКСПЕРИМЕНТ*
Уексперименті, проведеному в умовах теплиці, вивчені елементи продуктивності та біологічна фіксація азоту п'яти сортів люцерни: Полтавчанка, Віра, Лідія, Зайкевича (українські сорти) і Сітел (європейський сорт) в умовах водного стресу. Встановлено, що продуктивність люцерни лінійно падає з підвищенням водного дефіциту листків, а біологічна фіксація азоту люцерною зменшується з ростом дефіциту води аж до припинення. Не виявлена істотна сортова різниця в елементах продуктивності між вивченими сортами люцерни в умовах водного дефіциту. |
Постановка проблемы. Бобовые растения с присущей им симбиотической фиксацией атмосферного азота являются главным источником химически активного азота в материковых экологических системах. При этом адекватное снабжение водой поддерживает эффективное функционирование симбиозов всех бобовых культур. В засушливых системах земледелия использование азотных удобрений является рискованным из-за неустойчивых погодных условий, а использование летнего пара для сохранения влажности и снабжения минерализованным азотом производства зерна обычно приводит к повышенной деградации почвы (7). Известно, что во многих регионах бобовые культуры выращиваются в условиях недостатка воды, а это уменьшает накопление азота и урожайность (2-3, 6, 8, 12, 15). Проблема состоит в сохранении высокого уровня биологической фиксации азота в продуктивных сельскохозяйственных системах с помощью бобовых растений в условиях дефицита воды.
Анализ основных исследований и публикаций, в которых рассмотрено решение проблемы. Известно, что генетическая изменчивость бобовых культур в способности фиксировать азот в условиях засухи открыла возможность повышения устойчивости азотофиксации к недостатку влаги путем селекции (10, 15, 17). С другой стороны, известно, что клубенькообразование, рост и азотофиксация люцерны могут быть улучшены путем инокуляции растений конкурентоспособной и устойчивой к засухе культуре ризобия (6). Такая инокуляция может быть экономически оправданным методом увеличения продуктивности люцерны в условиях водного дефицита. Селекция растений и селекция ризобия – два пути увеличения устойчивости азотфиксирующих бобовых симбиотических систем в условиях водного дефицита. Существует также третий путь – обработка корней бобовых растений гормонами (2-3). Так, установлено, что обработка растений люцерны раствором ризобиального глюкана (0,06%) с последующей инокуляцией Sinorhizobium meliloti 441 ускоряло формирование мощного симбиотического аппарата (3). Такая обработка усиливала рост надземной биомассы люцерны как в условиях адекватного снабжения водой, так и в условиях водного дефицита. Показано, что обработка люцерны регуляторами роста ауксином и цитокинином на стадии бутонизации смягчало действие засухи на формирование и функционирование симбиотической системы с Sinorhizobium meliloti, стимулировала формирование клубеньков и фиксацию азота (2).
Интересны последние данные о чувствительности к засухе симбиотической фиксации азота ряда зернобобовых, включая сою, вигну, черный горошек, нут, бобы, люпин белый, горох и арахис (15-16). Так, относительная азотофиксация (ОАФ) на одно растение была определена для каждого вида как отношение величин азотофиксации индивидуальных водно-дефицитных растений и среднего значения азотфиксации хорошо обводненных растений. Далее этот показатель сравнивали с относительной скоростью транспирации (ОСТ). Результаты, полученные для сои, вигны и черного горошка, показали, что чувствительность ОАФ к почвенной засухе у них была выше, чем чувствительность ОСТ. Все остальные зернобобовые продемонстрировали ранее неизвестный эффект меньшей чувствительности ОАФ, по сравнению с ОСТ, в период водного дефицита. Авторы пришли к заключению о возможности селекции зернобобовых культур с пониженной чувствительностью симбиотической фиксации азота к дефициту воды для засушливых регионов. Так ли это для люцерны? Люцерна
______________
*Исследования выполнены в рамках международного научно-исследовательского проекта “BIO-Fix” (Австрия, Шри-Ланка, Украина), http://www. boku. ac. at/biofix/main. htm
имеет выдающиеся способности в области устойчивости к водному стрессу, хорошие адаптационные свойства (1), и ее селекцию продолжают проводить в Украине. В настоящее время известна продуктивность различных сортов люцерны, но ничего – о вкладе этой культуры в плодородие почв посредством биологической фиксации азота в условиях водного стресса. Исследователи и аграрии-практики нуждаются в такой информации для совершенствования систем севооборотов в неблагоприятных условиях окружающей среды, которые имеют тенденцию к ухудшению в связи с глобальным потеплением на планете.

Цель исследований и методика их проведения. Целью исследований являлось изучение способности различных сортов люцерны первого года вегетации фиксировать атмосферный азот в условиях водного стресса. Для исследования были сформулированы следующие направления:
1. Изучить способность пяти сортов люцерны фиксировать атмосферный азот при различных уровнях (три уровня) снабжения водой.
2. Изучить продуктивность надземной и подземной частей пяти сортов люцерны в условиях разного уровня (три уровня) снабжения водой.
3. Определить сортовую разницу в фиксации атмосферного азота у различных сортов люцерны?
В работе использовали семена следующих сортов люцерны, представленные Полтавским институтом АПВ им. УААН: Полтавчанка (национальный стандарт), Вира, Лидия, Зайкевича и европейский сорт Sitel (Сител), представленный профессором Б. Фрейером из института органического земледелия аграрного университета BOKU (Вена, Австрия). В качестве культуры сравнения использовали кострец безостый (Bromopsis inermis (L.) Colub), который выращивали одновременно в одинаковых с люцерной условиях, и который не способен фиксировать атмосферный азот. Семена высевали в пластиковые сосуды диаметром 20 см и высотой 50 см, заполненные одинаковым количеством тщательно перемешанного грунта из 30-ти см слоя полевых посевов люцерны. Количество повторностей – четыре. Вегетационные сосуды размещали на воздухе в домики с пленочным укрытием от дождя. Количество домиков – три и соответственно три режима снабжения водой. Схемы расположения сосудов в каждом домике была рендомизованы. Водные режимы создавали дозированным поливом и контролировали измерением водного дефицита листьев перед каждым поливом. Семена в сосуды высевали нормой 40 кг/га (0,12 г на сосуд) и давали развиваться всходам две недели с хорошим увлажнением (поверхность почвы была постоянно влажной). Через две недели после всходов оставляли по 20 растений на сосуд и начинали ограничивать полив в двух из трех домиков. В одном из вариантов полив проводили таким образом, чтобы поверхность почвы была постоянно влажной. В двух других вариантах доводили засуху до появления признаков увядания растений, восстанавливая тургор разными количествами воды. Самым дефицитным по воде был домик №2, самым увлажняемым – №1. Таким образом были созданы три режима водоснабжения люцерны и культуры сравнения – костреца безостого. Перед цветением растения извлекали из контейнеров, измеряли, определяли сухую массу побегов и корней (+5 см стебля над поверхностью почвы) путем высушивания при 65°C до постоянной массы. Далее анализировали растительные образцы на содержание азота по Кьельдалю, определяя величину биологической азотфиксации (BNF – biological nitrogen fixation) дифференциальным методом (13) без учета почвенного минерального азота по формуле:
BNF [кг/гa] = (поб. Nлюц. + кор. Nлюц.) [кг/га] –(поб. Nкостр. + кор. Nкостр.) [кг/гa],
где: (поб. Nлюц. + кор. Nлюц.) : содержание азота в побегах и корнях люцерны кг/га;
(поб. Nкостр. + кор. Nкостр.): содержание азота в побегах и корнях костреца, кг/га.
Относительное содержание воды в листьях люцерны Rl определяли по формуле:
Rl = (свежий вес – сухой вес)/(тургорный вес – сухой вес).
Относительный водный дефицит WSD (water saturation deficit) определяли как разницу 1-Rl. Определение WSD проводили каждые 4-5 дней после начала засухи следующим образом: срезали по три листа с каждой повторности, взвешивали (это свежий вес) и помещали в дистиллированную воду на время, достаточное для полного насыщения листьев водой, затем удаляли фильтровальной бумагой остатки воды с поверхности листьев и снова взвешивали (это тургорный вес). Время полного насыщения листьев люцерны водой определяли в отдельном эксперименте, взвешивая листья через двухчасовые промежутки до получения максимальной и не изменяющейся по времени массы. Для определения сухого веса тургорные листья высушивали при 65°С до постоянной массы.
Водный потенциал листьев люцерны определяли с помощью камеры давления (Plant Water Status Consol 3000, Soilmoisture Equipment Corp. USA) в соответствии с процедурой, описанной Брауном и Таннером (8-9, 11). Измерения проводили по достижению растениями высоты 15 см с одним листом из каждой повторности после 17 часов.
Индекс листовой поверхности по завершению эксперимента (LAI – leaf area index) рассчитывали как площадь в м2 общей листовой поверхности люцерны, приходящейся на 1 м2 поверхности почвы. Вначале измеряли поверхность нескольких листьев, определяли их сухую массу и рассчитывали площадь поверхности 1 г сухой массы листа. Собирали все листья каждой повторности и определяли их сухую массу, которую пересчитывали в общую поверхность аналогично, используя величину поверхности 1 г сухой массы (4).
Результаты исследований. Следствием водного стресса в домиках №2 и №3 явилось снижение массы побегов и корней всех растений (рис. 1, 2) в сравнении с вариантом достаточного увлажнения в домике №1, естественно, снижался и индекс листовой поверхности (рис. 3). Однако не было обнаружено существенных различий между сортами в реакции на водный стресс. У всех сортов снижение продуктивности хорошо коррелировало с общей суммой водного дефицита растений за восемь измерений в процессе вегетации. График корреляций представлен на рис. 4.
Водный потенциал растений изменялся от 5 до 19 Бар. Этот показатель тем меньше, чем лучше растение было снабжено влагой. Ввиду регулярных осадков, колебания водных потенциалов в полевых измерениях тех же сортов люцерны первого года вегетации составляли 1,5-8,5 Бар. Проведенные измерения, тем не менее, не обнаружили существенной разницы между сортами, вероятно, по причинам: а) высокой вариабельности развития корневой системы между индивидуальными растениями в первый год роста (8-9), и б) невозможностью измерения водного потенциала с точностью большей, чем 0,1 Бар. Кроме того, водный потенциал растений может меняться в значительных пределах в течение дня в зависимости от температуры и влажности воздуха. Так, при измерении водных потенциалов люцерны в полевых посевах, при достаточном увлажнении из-за большого количества осадков в период измерений, наблюдали колебания значений водного потенциала от 7 Бар в 15 часов 40 минут до 3 бар в 17 часов 45 минут.
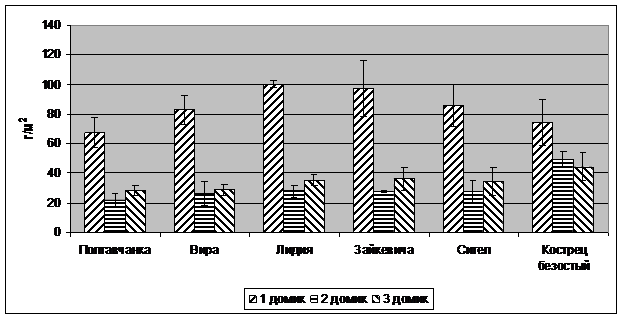
Рис. 1. Сухая масса побегов люцерны и костреца
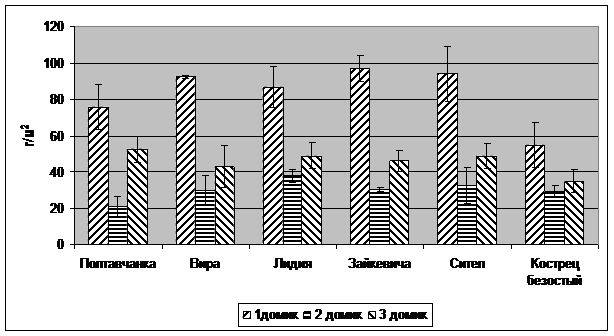
Рис. 2. Сухая масса корней люцерны и костреца
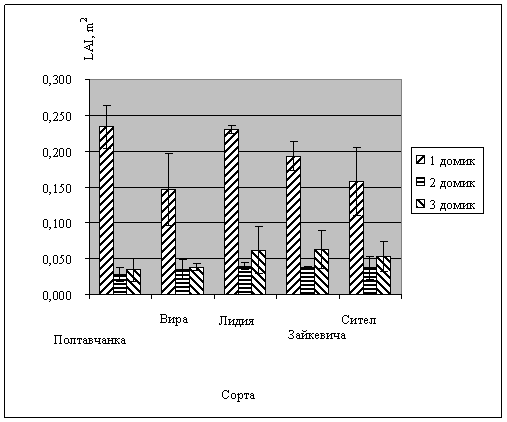

Рис. 3. Индекс листовой поверхности люцерны в зависимости от уровня снабжения водой
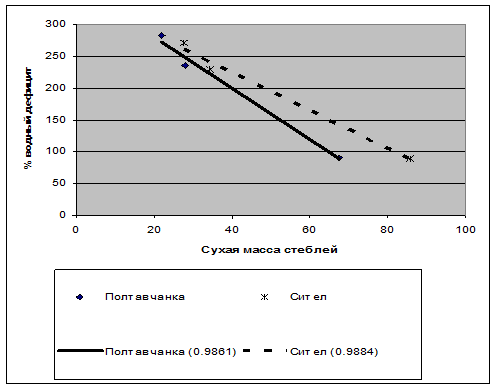
Рис. 4. Корреляция продуктивности люцерны с водным дефицитом листьев (сумма водных дефицитов в % за восемь измерений в процессе вегетации). Коэффициенты корреляции составили от 0,8631 до 0,9981.
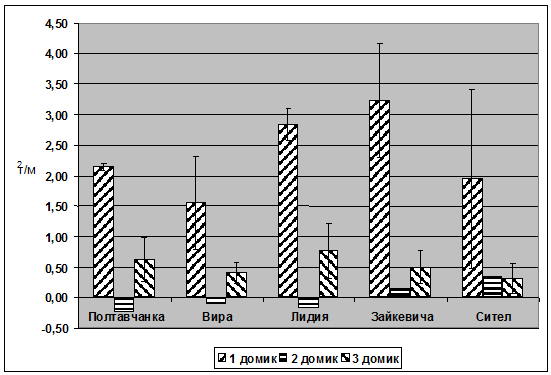
Рис. 5. Величина фиксации азота сортами люцерны при разных уровнях снабжения водой
Следствием водного стресса в эксперименте явилось резкое снижение биологической фиксации азота люцерной с 4,2 г/м2 практически до нуля в вариантах с минимальным увлажнением. Отрицательные значения азотофиксации засушенных растений люцерны демонстрируют факт лучшего использования минерального почвенного азота кострецом в стрессовых условиях эксперимента. Как и в случае с другими вышеприведенными показателями, не обнаружено существенной сортовой разницы в фиксации атмосферного азота люцерной в условиях водного стресса.
Выводы: 1. Продуктивность люцерны линейно падает с повышением водного дефицита листьев.
2. Биологическая фиксация азота люцерной уменьшается с ростом дефицита воды вплоть до прекращения.
3. Не обнаружена существенная сортовая разница в элементах продуктивности между изученными сортами люцерны в условиях водного дефицита.
БИБЛИОГРАФИЯ
1. Зінченко Б. С. Люцерна і клевер. – К.: Урожай, 1989. – 232 с.
2. , , Регуляция азотофиксации и фотосинтеза у люцерны с помощью цитокининов и ауксинов в различных водных условиях// Физиол. и биохим. культ. раст. – 2002. – Т.34. – №5. – С.317-325.
3. , , и др. Влияние глюкана Sinorhizobium meliloti на формирование и функционирование азотфиксирующих симбиотических систем у люцерны при различных уровнях снабжения водой// Физиол. и биохим. культ. раст. – 2002. – Т.34. – №5. – С. 413-418.
4. Asner G. P., Scurlock J. M.O. and Hicke J. A. Global synthesis of leaf area index observations: implications for ecological and remote sensing studies// Global Ecology&Biogeography. – 2003. – Vol. 12. – P. 191-205.
5. Athar M., Johnson D. A. Nodulation, biomass production, and nitrogen fixation in alfalfa under drought// Journal of Plant Nutrition. – 1996. – P. 185-199.
6. Athar, Mohammad; Johnson, Douglas A. Influence of drought on competition between selected Rhizobium meliloti strains and naturalized soil rhizobia in alfalfa// Plant and Soil/ – 1996. – Vol. 184. – No 2. – P. 231-241.
7. Biederbeck V. O., Bouman O. T., Campbell C. A., et al. Nitrogen benefits from four green-manure legumes in dryland cropping systems// Canadian Journal of Plant Science. – 1996. – Vol. 76. – P. 307-315.
8. Brown P. W., Tanner C. B. Alfalfa Water Potential measurement: A Comparison of the Pressure Chamber and Leaf Dew-point Hygrometers// Crop Science/ – 1981. – Vol. 21. – P. 240-244.
9. Brown P. W., Tanner C. B. Alfalfa Stem and Leaf Growth during Water Stress// Crop Science. – 1983. – Vol. 75. – P. 799-805.10. Graham P. H., Vance C. P. Nitrogen fixation in perspective: an overview of research and extension needs// Field Crops Research. – 2000. – Vol. 65, 93-106.
11. Internet resources. http://www. /~pms/home. htm http://weather. nmsv. edu/Teaching_Material/Soil698/Student_Material/pressurebomb/bomb. htm
12. Kurdali, Fawaz; Al-Ain, Farid; Al-Shamma, Mohammed. Nodulation, dry matter production, and N2 fixation by fababean and chickpea as affected by soil moisture and potassium fertilizer// Journal of Plant Nutrition.- 2002. – Vol. 25. – N2/ – P. 355-368.
13. Munro J. M.M., Davis D. A. Potential pasture production in the uplands of Wales.5.The nitrogen contribution of white clover// Journal of the British Grassland Society. – 1974. – Vol. 29. – P. 213-223.
14. O`Hara G. W. Nutritional constraints on root nodule bacteria affecting symbiotic nitrogen fixation: a review// Australian Journal of Experimental Agriculture. – 2001. – Vol. 41. – P. 417-433.
15. Serraj, Rachid; Sinclair, Thomas R.; Purcell, Larry C. Symbiotic N2 fixation response to drought// Journal of Experimental Botany/ – 1999/ – Vol. 50/ – No 331. – P. 143-155.
16. Sinclair T. R., Serraj R. Dinitrogen Fixation Sensitivity to Drought among Grain Legume Species// Nature. – 1995. – No 378. – P. 344.
17. Zahran H. H. Rhizobium-Legume Symbiosis and Nitrogen Fixation under Sever conditions and in an Arid Climate// Microbiology and Molecular Biology reviews. – 1999. – Vol. 63. – No.4. – P. 968-989.
