
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Истощение запасов креатинфосфата
Вспомним, что креатинфосфат используется в анаэробных условиях для восстановления энергобогатой АТФ по мере ее расходования, следо
вательно, для поддержания запасов АТФ в организме. Биопсические исследования мышц бедер человека показали, что во время повторяющихся максимальных сокращений мышц возникновение утомления совпадает с истощением запасов КФ. Хотя подобные виды деятельности обеспечиваются энергией АТФ, ее запасы во время мышечных усилий истощаются не так быстро, как запасы КФ, поскольку в образовании АТФ принимают участие и другие системы. Однако после истощения запасов КФ способность организма быстро восполнять запасы использованной АТФ серьезно нарушается. Использование АТФ продолжается, но возможность системы АТФ —КФ восполнять расходуемую АТФ значительно уменьшается. В результате уровни АТФ также понижаются. В состоянии изнеможения могут быть истощены запасы как КФ, так и АТФ.
Чтобы задержать возникновение утомления, спортсмену необходимо контролировать интенсивность усилий исходя из наиболее оптимальной скорости выполнения физического упражнения, чтобы не допустить преждевременного истощения запасов КФ и АТФ. Если начальная скорость очень высока, запасы АТФ и КФ быстро уменьшатся, что приведет к раннему утомлению и неспособности сохранить скорость на заключительных стадиях дистанции. Тренировка и опыт позволяют спортсмену находить оптимальную скорость, при которой обеспечивается наиболее эффективное использование АТФ и КФ в течение всей мышечной деятельности.
Истощение запасов гликогена
Поддержание уровней АТФ обеспечивается также аэробным и анаэробным расщеплением мышечного гликогена. В видах спорта, в которых интенсивные мышечные усилия длятся всего несколько секунд, мышечный гликоген является основным источником энергии для синтеза АТФ. К сожалению, его запасы ограничены и быстро истощаются.
Как и в случае с КФ, скорость истощения запасов мышечного гликогена зависит от интенсивности физической нагрузки. Увеличение интенсивности приводит к снижению уровня мышечного гликогена. Во время бега на спринтерские дистанции, например, мышечный гликоген расходуется в 35 — 40 раз быстрее, чем при ходьбе. Наличие мышечного гликогена может быть ограничивающим фактором даже при средних усилиях. Для удовлетворения высоких потребностей мышцы в энергии во время физической нагрузки необходимо постоянное поступление гликогена.
Как видно из рис. 5.17, мышечный гликоген быстрее используется в первые минуты физической деятельности. Несмотря на то, что испытуемый бежал в одном темпе, интенсивность исполь-
105
зования гликогена в икроножной мышце была максимальной в первые 90 мин. Испытуемый сообщал об испытываемом напряжении (как трудно ему давалось усилие) в различные моменты теста. Он испытывал среднюю степень стресса в самом начале, когда запасы гликогена были еще достаточно большими, несмотря на высокую интенсивность его использования. Он не испытывал значи
тельного утомления до тех пор, пока запасы мышечного гликогена почти не истощились. Таким образом, возникновение ощущения утомления при продолжительной физической нагрузке совпадает со снижением уровня мышечного гликогена. Марафонцы называют внезапное наступление утомления на 29— 35-м километре дистанции (18— 22-я миля) "столкновением со стенкой". Частично возникновение чувства утомления обусловлено истощением запасов мышечного гликогена.
Истощение запасов гликогена в различных мышечных волокнах. Существуют определенные паттерны рекруитирования вовлечения мышечных волокон и истощения в них запасов гликогена. Запасы гликогена могут истощиться в наиболее часто используемых волокнах. Это ведет к сокращению числа волокон, способных производить мышечное усилие, необходимое для выполнения физического упражнения.
На рис. 5.18 представлена микроструктура мышечных волокон, взятых из мышцы бегуна до и после забега на 30 км. На рис. 5.18,а мы видим дифференциацию МС- и БС-волокон. Одно из БС-волокон обведено кругом. На рис. 5.18,^продемонстрирован второй образец той же мышцы, окрашенной для выявления гликогена. Чем краснее (темнее) окраска, тем больше в ней гликогена. До забега все волокна содержали достаточное количество гликогена и имели красную окраску. В обведенном круге БС-волокне на рис. 5.18,6 (после забега) все еще достаточно гликогена. В то же время в находящихся рядом с ним МС-волок-нах запасы гликогена почти полностью истощены. Это свидетельствует о том, что МС-волокна более интенсивно используются во время мышечной деятельности, требующей проявления выносливости, во время которой производится среднее усилие, например, бег на 30 км.
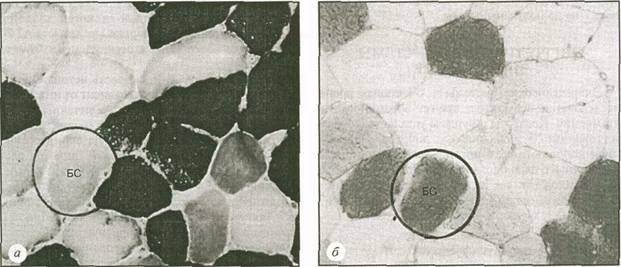
Рис. 5.18. Гистохимическое окрашивание на выявление мышечного гликогена до (а) и после (б)
забега на 30 км. Пояснения в тексте
106
Паттерн истощения запасов гликогена в МС-и БС-волокнах зависит от интенсивности физической нагрузки. Вспомним, что МС-волокна первыми вовлекаются при невысокой интенсивности физической нагрузки. По мере увеличения интенсивности начинают включаться и БСд-во-локна. Когда интенсивность возрастает до максимальной, наступает очередь БСц-волокон. По-видимому, такой же паттерн характерен и для процесса истощения запасов гликогена.
На рис. 5.19 показано количество гликогена, который использовали МС-, а также БС^- и БСд-волокна латеральной широкой мышцы бедра при езде на велосипеде с различным процентом МПК испытуемого. При относительно невысокой интенсивности физической нагрузки (40 — 60 % МПК) наиболее активными были МС-волокна. При более высокой интенсивности (75 — 90 % МПК) чаще использовались БС-волокна, которые быстрее истощали свои запасы гликогена по сравнению с МС-волокнами.
Это, однако, не означает, что МС-волокна меньше используются во время максимальных сокращений, чем БС, просто здесь отражен тот факт, что БС-волокна в большей степени "полагаются" на гликоген. При интенсивных мышечных сокращениях в работе участвуют все типы волокон.
Когда в МС-волокнах истощаются запасы гликогена, БС-волокна оказываются либо неспособными производить достаточное усилие, либо не могут быть полностью рекруитированы, чтобы компенсировать недостаточное усилие. Поэтому выдвигалось предположение, что ощущения мышечного утомления или тяжести при продолжительной физической нагрузке могут отражать неспособность некоторых мышечных волокон реагировать на требования, предъявляемые физической нагрузкой.
Истощение запасов гликогена в различных мышечных группах. Помимо избирательного истоще-
ния запасов гликогена в МС - и БС-волокнах, физические нагрузки могут предъявлять чрезмерно высокие требования к отдельным мышечным группам. Рассмотрим исследование, в котором испытуемые бегали на тредбане, установленном в трех положениях — горизонтально, с наклоном вниз и с наклоном вверх, — в течение 2 ч при 70 % МПК. Как показывают результаты (рис. 5.20), независимо от того, бежит ли спортсмен по наклонной вниз, вверх или горизонтально, больше всего гликогена использует икроножная мышца. Это свидетельствует о том, что мышцы-разгибатели голеностопного сустава, подвергавшиеся наибольшему физическому стрессу во время бега на длинные дистанции, скорее всего истощают запасы гликогена в отличие от мышц бедер. Таким образом, участком возникновения утомления являются мышцы нижней части ног.
Истощение запасов гликогена и глюкоза крови
Один лишь мышечный гликоген не в состоянии обеспечить достаточное количество углеводов при физических нагрузках продолжительностью несколько часов. Значительное количество энергии во время физических нагрузок, требующих проявления выносливости, обеспечивает глю-
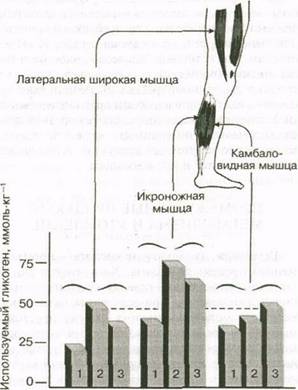
Рис. 5.20. Использование запасов гликогена икроножной, камбаловидной и латеральной широкой мышцами во время бега по прямой (1), под уклоном вверх (2) и вниз (3)
107
коза, которая поступает в мышцы с кровью. Запасы гликогена в печени расщепляются, что обеспечивает постоянное поступление глюкозы в кровь. В первые моменты выполнения упражнения для образования энергии необходимо относительно немного глюкозы крови, однако позднее вклад глюкозы в образование энергии значительно увеличивается. Чтобы обеспечить адекватное потребление глюкозы мышцами, по мере увеличения продолжительности физической нагрузки печени приходится расщеплять все больше и больше гликогена.
Запасы гликогена в печени ограничены, и она не может быстро расщеплять глюкозу из других субстанций. Следовательно, когда потребление мышцами глюкозы превышает образование ее печенью, уровни глюкозы крови понижаются. Не имея возможности получить достаточное количество глюкозы из крови, мышцы вынуждены более интенсивно использовать свои резервы гликогена, что ускоряет истощение его запасов в мышцах и приводит к раннему возникновению утомления.
Влияние гликогена на мышечную деятельность
Не вызывает удивления тот факт, что увеличение снабжения мышц гликогеном в начале физической нагрузки на выносливость улучшает результаты. Значение накопления мышечного гликогена для мышечной деятельности, требующей проявления выносливости, обсуждается в главе 15. Пока отметим, что истощение запасов гликогена и гипогликемия (пониженное содержание глюкозы в крови) ограничивают уровень мышечной деятельности в видах спорта аэробной производительности. Утомление в видах спорта более короткой продолжительности, по-видимому, — результат аккумуляции промежуточных продуктов метаболизма, таких, как лактат и Н^ в мышцах.
ПРОМЕЖУТОЧНЫЕ ПРОДУКТЫ МЕТАБОЛИЗМА И УТОМЛЕНИЕ
Вспомним, что молочная кислота — промежуточный продукт гликолиза. Хотя многие считают, что именно она — главная причина утомления и изнеможения при всех видах физических нагрузок, молочная кислота аккумулируется в мышечном волокне только во время кратковременного мышечного усилия высокой интенсивности. У марафонцев, например, уровни молочной кислоты в конце забега могут быть почти такими, как в состоянии покоя, несмотря на крайнюю степень утомления. Как уже отмечалось, возникновение утомления у марафонцев обусловлено недостаточным энергетическим обеспечением, а не избытком молочной кислоты.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 |
