


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Тем же ферментом (это очень важно) «вырезают» нужный ген (или участок ДНК, содержащий этот нужный – целевой ген) из какого-то другого плазмидного вектора. Если этот нужный участок не очень велик, то его можно синтезировать (что конечно, более затратно), оснастив соответствующими «концами» (окончаниями) с обеих сторон).
Так как действовали в обоих случаях одними и теми же ферментами (т. е. ферментами, узнающими и расщепляющими одни и тем же специфические нуклеотидные последовательности (сайты) в двуцепочечной молекуле ДНК, то выполняется важное в молекулярном клонировании обстоятельство о совпадении «концов» принимающей информацию молекулы ДНК и внедряемого фрагмента ДНК.
Гидролиз («расщепление») молекулы ДНК в нужном сайте эндонуклеазами рестрикции (рестриктазами) «липких» концов или (прямых) «тупых» концов.
2 этап – встраивание фрагментов в принимающую ДНК и лигирование. Проводится совместное выдерживание «разрезанной» плазмидной ДНК и участка, содержащего интересующий исследователя ген в течение определенного времени в необходимых термальных условиях. В результате этого становится возможным объединение плазмиды и гена при помощи фермента ДНК-лигазы, который их как бы «сшивает», образуя ковалентные фосфодиэфирные связи. Возникает новая рекомбинантная плазмидная ДНК (новый вектор).
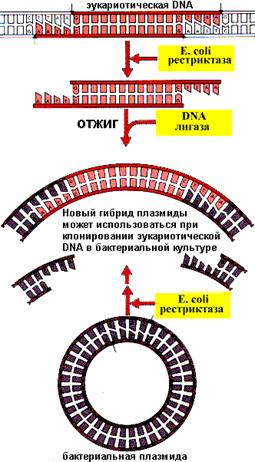 Полученный новый вектор выдерживается в течение определенного времени совместно с микроорганизмами-реципиентами. В целом ряде случаев плазмида-вектор внедряется внутрь клеток. Этот процесс носит название – трансформация. Для отбора трансформированных клеток смесь (протрансформированных и непротрансформированных) микроорганизмов высевают на среду, содержащую соответствующий фактор селективности (например, тот же антибиотик, ген устойчивости к которому был в плазмиде). Выросшие на селективной среде колонии проверют на способность к синтезу нужного биологического объекта.
Полученный новый вектор выдерживается в течение определенного времени совместно с микроорганизмами-реципиентами. В целом ряде случаев плазмида-вектор внедряется внутрь клеток. Этот процесс носит название – трансформация. Для отбора трансформированных клеток смесь (протрансформированных и непротрансформированных) микроорганизмов высевают на среду, содержащую соответствующий фактор селективности (например, тот же антибиотик, ген устойчивости к которому был в плазмиде). Выросшие на селективной среде колонии проверют на способность к синтезу нужного биологического объекта.
Процесс разделения геномной ДНК на клонируемые элементы и введения этих элементов в клетки-хозяева называется созданием геномной библиотеки (банка клонов, банка генов).
Все системы клонирования должны отвечать двум основным требованиям:
1) наличию нескольких сайтов для клонирования;
2)возможности достаточно простой идентификации клеток с рекомбинантными ДНК.

Для всех рутинных процедур молекулярного клонирования широко используется E. coli в качестве клетки-хозяина. Клетки, способные поглощать чужеродную ДНК, называются компетентными. Компетентность E. coli повышают, используя специальные условия культивирования. Для получения больших количеств чужеродных белков с помощью рекомбинантных штаммов E. coli была сконструирована плазмида, содержащая сильный промотор, селективный маркерный ген и короткий участок с несколькими уникальными сайтами для рестрицирующих ферментов – полилинкер.
Эффективными методами трансформации E. coli плазмидами является электропорация (воздействие на клеточные мембраны электрическим током для увеличения их проницаемости). Для введения клонированных генов в соматические клетки также применяют микроинъекции и микроукалывания или слияние с клеткой нагруженных ДНК мембранных везикул (липосом).
Получение рекомбинантного инсулина
• Инсулин - пептидный гормон, выделяемый β-клетками о. Лангенгарса.
• Состоит из двух пептидных цепей: А-цепь - из 21 аминокислотных остатков. В-цепь содержит 30 аминокислотных остатков
• Эти две цепи связаны бисульфидными –S-S - связями, которые обеспечивают пространственную структуру белка инсулина.
• При синтезе инсулина в поджелудочной железе вначале образуется предшественник инсулина - проинсулин.
• Проинсулин состоит из А-цепи, В-цепи и С-пептида, состоящего из 35 аминокислотных остатков.
• С-пептид отщепляется под действием карбоксипептидазы и трипсина и проинсулин переходит в активный инсулин.
• До получения рекомбинантного инсулина препарат получали из поджелудочной железы свиней и крупного рогатого скота
Инсулин был первым лекарственным рекомбинантным препаратом, полученным в промышленных масштабах еще в 1982 г. Как известно, инсулин широко применяется при лечении инсулино-зависимой формы сахарного диабета.
До получения рекомбинантного инсулина препарат получали из поджелудочной железы свиней и крупного рогатого скота. Однако такой способ получения инсулина имел целый ряд недостатков:
1)недостаток поголовья скота;
2)сложности хранения и транспортировки сырья;
3)трудности выделения и очистки гормона;
4)возможность развития аллергических реакций.
В настоящее время инсулин человека, в основном, получают двумя способами:
1) модификацией свиного инсулина синтетико-ферментативным методом;
Метод основан на том, что свиной инсулин отличается от инсулина человека одной заменой на С-конце В-цепи Ala30Thr. Замену аланина на треонин осуществляют путем катализируемого ферментом отщепления аланина и присоединение вместо него защищенного по карбоксильной группе остатка треонина, присутствующего в реакционной смеси в большом избытке. После отщепления защитной О-трет-бутильной группы получают инсулин человека.
2) генно-инженерным способом;
Существует два основных подхода для получения генно-инженерного инсулина человека.
В первом случае (2.1) осуществляют раздельное (разные штаммы-продуценты) получение обеих цепей с последующим фолдингом молекулы (образование дисульфидных мостиков) и разделением изоформ.
Во втором (2.2) - получение в виде предшественника (проинсулина) с последующим ферментативным расщеплением трипсином и карбоксипептидазой В до активной формы гормона.
Наиболее предпочтительным в настоящее время является получение инсулина в виде предшественника, обеспечивающее правильность замыкания дисульфидных мостиков (в случае раздельного получения цепей проводят последовательные циклы денатурации, разделения изоформ и ренатурации).
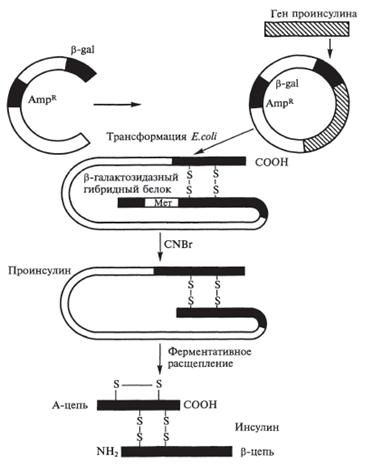
Метод 2.1. Раздельный синтез А - и В-цепей с последующим заключением между ними дисульфидных связей.
• 1. Путем химического синтеза создаются последовательности нуклеотидов, которые кодируют образование А и В цепей (создание синтетических генов).
• 2. Каждый из синтетических генов вводят в плазмиды (в одну плазмиду вводят ген, синтезирующий цепь А, в другую плазмиду вводят ген, синтезирующий цепь В).
• 3. Вводят ген, кодирующий образование фермента бетагалактозидазы. Этот ген включают в каждую плазмиду для того, чтобы добиться активной репликации плазмид.
• 4. Плазмиды вводят в клетку E. coli - кишечной палочки и получают две культуры продуцента, одна культура синтезирует А-цепь, вторая В-цепь.
• 5. Помещают две культуры в ферментер. В среду добавляют галактозу, которая индуцирует образование фермента бетагалактозидазы. При этом плазмиды активно реплицируются, образуя много копий плазмид и, следовательно, много генов, синтезирующих Аи В цепи.
• 6. Клетки лизируют, выделяют А и В цепи, которые связаны с бетагалактозидазой. Все это обрабатывают бромцианом и отщепляют А и В-цепи от бетагалактозидазы. Затем производят дальнейшую очистку и выделение А и В цепей.
• 7. Окисляют остатки цистеина, связывают и получают инсулин.

Недостатки подобного метода: надо получать два отдельных штамма-продуцента, проводить две ферментации, две процедуры выделения и очистки, а самое главное, трудно обеспечить правильное замыкание дисульфидных связей, то есть получить активный инсулин.
Метод 2.2. Синтез проинсулина с последующим выщеплением С-_пептида.
При этом конформация проинсулина обеспечивает правильное замыкание дисульфидных связей, что делает второй способ микробиологического синтеза более перспективным.

В Институте биоорганической химии РАН получен рекомбинантный инсулин (инсуран) с использованием генно-инженерных штаммов E. coli. Из выращенной биомассы выделяется предшественник, гибридный белок, экспрессируемый в количестве 40% от всего клеточного белка, содержащий препроинсулин. Превращение его в инсулин in vitro осуществляется в той же последовательности, что и in vivо – отщепляется лидирующий полипептид, препроинсулин превращается в инсулин через стадии окислительного сульфитолиза с последующим восстановительным замыканием трех дисульфидных связей и ферментативным вычленением связывающего С-пептида. После ряда хромотографических очисток, включающих ионообменные, гелевые и ВЭЖХ, получают человеческий инсулин высокой чистоты и природной активности.
С-Пептид проинсулина человека был описан в 60-х годах XX века. Первоначальные исследования не обнаружили существование у него специфической биологической функции. Долгое время считалось, что он необходим только для образования правильной конформации белковой молекулы инсулина. В начале 90-х годов XX века появилось много сведений о применении с-пептида для профилактики и лечения осложнений, связанных с диабетом. с-Пептид с успехом применялся для улучшения состояния пациентов, страдающих от ретинопатии, ангиопатии и нефропатии - основных факторов инвалидизации и смертности при диабете. Однако в настоящее время крупномасштабные исследования и применение в клиниках затруднены из-за недостатка с-пептида на рынке, его высокой стоимости и низкого качества. Так, например, известный поставщик реактивов - корпорация Sigma-Aldrich - предлагает с-пептид, имеющий чистоту 85% по цене 371,5 евро за 250 мкг препарата. Другая корпорация - Phoenix Pharmaceuticals, Inc. - предлагает 200 мкг препарата по цене 75,00 долларов.
В отличие от инсулина аминокислотная последовательность с-пептида сильно отличается у разных видов млекопитающих, что делает невозможным получение его из животного сырья. Существующие способы получения с-пептида, можно разделить на три категории:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
