
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Характерным признаком морозостойкости растений, в том числе и сосны обыкновенной, является продолжение жизнедеятельности при отрицательной температуре, хотя и резко ослабленном виде. Способность к ассимиляции СО2 при фотосинтезе у сосны сохраняется при снижении температуры до -50 С. . Дыхание продолжается и при более низких температурах, вплоть до -200С. В зимний период, прежде чем дыхательный процесс полностью не инактивируется, возможен переход растений на анаэробный путь дыхания, менее чувствительный к повреждающему действию низких температур [12].
Даже слабо протекающий фотосинтез и дыхание позволяют сосне поддерживать более высокую внутриклеточную температуру по сравнению с температурой окружающей среды. При сравнении температуры ткани хвои и окружающего воздуха в первой декаде апреля отмечено, что при полном солнце и слабом ветре температура хвои превышала температуру воздуха на 12–140 С [8 ].
Адаптация к крайним условиям существования предполагает компенсаторные изменения обмена веществ, позволяющие поддерживать метаболические реакции на определенном уровне, обеспечивающем выполнение основных жизненных функций. Одним из наиболее широко распространенных способов низкотемпературной адаптации является изменение свойств ферментов, определяющих скорость и направленность метаболических процессов в организме. Следует отметить, что изменение активности и изоэнзимного спектра ферментов является общей ответной реакцией организма на любое стрессовое воздействие и поэтому неспецифично к холоду.
В целом изучение механизма низкотемпературной адаптации у различных климатипов сосны обыкновенной позволяет лучше понять особенности ее метаболизма в различных частях ареала и выявить физиологические и биохимические признаки устойчивости сосны к низкотемпературному фактору среды.
1.3. Физиологическое значение марганца
Наиболее широко распространенным способом адаптации к низким отрицательным температурам является изменение активности белков-ферментов. Хорошо известно, что в состав белков-ферментов входят микроэлементы. Среди них большое значение имеет марганец. Роль марганца и пигментов фотосинтеза в адаптации хвои сосны обыкновенной к низким температурам может проявляться как прямо, так и косвенно. Марганец входит в состав ферментов, которые катализируют синтез низкопротекторных соединений (свободной аминокислоты, полиаминов), играющих исключительно важную роль в морозоустойчивости растений. В этом выражается его опосредованное влияние. Прямое влияние марганца на процесс адаптации заключается в том, что он входит в состав флавиновых ферментов, участвующих в синтезе порфобиллиногена, а также участвует в упрочнении связи хлорофилл–белок, предохраняя пигмент от преждевременного окисления.
Большое значение для внутриклеточного обмена веществ имеют соединения марганца с ферментами, обуславливающие усиление окислительно-восстановительных процессов в растениях. Марганец благодаря своей способности к перемене валентности при окислении-восстановлении имеет существенное значение в процессе фотосинтеза. Положительное влияние этого элемента на фотосинтез показал ещё Мак-Арг (1922). Этот микроэлемент элемент способен регулировать направленность окислительно-восстановительных процессов в растениях в зависимости от различных условий питания: при аммиачном питании он действует как сильный окислитель, при нитратном питании – как восстановитель [13]. Так как фотосинтез является процессом окислительно-восстановительным, а марганец способен регулировать окислительно-восстановительные процессы, можно было предположить, что он будет оказывать существенное влияние на интенсивность фотосинтеза.
Марганец значительно повышает скорость потребления кислорода суспензией хлоропластов не только на свету, но и в темноте. Дефицит марганца (и меди) в растениях ведет к уменьшению содержания в них не только хлорофилла, но каротина и ксантофила. Из клеточных органелл больше всего марганца содержится в хлоропластах, около 30% всего количества марганца, находящегося в клетке [14; 15]. Весь марганец хлоропластов находится в прочносвязанном с белком состоянии, образуя соединения хелатного типа.
2. Экспериментальная часть
2.1. Цель и задачи исследования
Цель работы: изучить динамику содержания марганца и пигментов фотосинтеза в хвое сосны обыкновенной в зависимости от сезонности.
Задачи:
1. Выявить динамику содержание основных и дополнительных пигментов в хвое сосны обыкновенной в зависимости от сезонности.
2. Определить содержание марганца в хвое сосны обыкновенной фотометрическим методом в зависимости от сезонности.
3. Провести статистическую обработку полученных данных, результаты свести в таблицы и оформить графически.
4. На основании полученных результатов и теоретического материала предположить механизмы адаптации хвойных растений к действию неблагоприятных факторов внешней среды.
2.2. Объекты и методы исследования
Объектами исследования служили хвоинки сосны обыкновенной (Pinus silvestris L.).
Pinus silvestris L.- сосна обыкновенная - дерево 20-40 м высоты и 80-100 см в диаметре. Ствол в насаждении прямой, высоко очищенный от сучьев, на открытом месте сбежистый, ветвистый, с кроной, начинающейся почти у земли. Кора снизу ствола красно-бурая, глубоко-бороздчатая, в верхней части и на старых ветвях желтовато-красная, почти гладкая, на молодых ветвях серо-зеленая. Естественно произрастает почти по всей Европе (от Испании и Греции до Заполярья), в Сибири. Встречается в различных экологических условиях - на бедных и сухих песчаных почвах, на каменистых породах в горах, на торфяно-болотных почвах. Быстрорастущая древесная порода. Светолюбива и морозостойка.
Образцы хвои первого и второго года отбирали из средней части кроны 20-25-летних деревьев. Содержание основных и дополнительных пигментов определяли с помощью спектрофотометра СФ-26 по методике описанной в спиртовых растворах пигментов (этанол 96%). [16 ].
Концентрацию пигментов рассчитывали по формулам (Wintermans, De Mots, 1965):
а) для хлорофиллов
СА= 13,70*Д665 - 5,76*Д649
СВ= 25,80*Д649 - 7,66*Д665
СА+В = 6,10*Д665 + 20,04*Д649
б) для каротиноидов (Веттштейн,1949)
Скар= 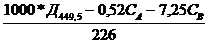
Содержание пигментов в растительном материале в мг/г сырого веса рассчитывали по формуле:
А = 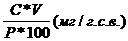 , где
, где
С - концентрация пигмента в мг/л;
V - объем вытяжки пигмента в мл;
Р - навеска растительного материала в г;
А - содержание пигмента в растительном материале в мг/г сырого веса.
Содержание марганца определялось фотометрическим методом, описанным [13 .
Концентрацию полученного раствора вычисляли по методу сравнения оптической плотности стандартного и исследуемого растворов по уравнению:
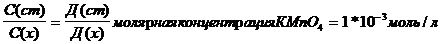
Массу марганца в мг/кг абсолютно сухого вещества вычисляли по формуле:
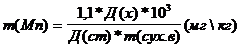
Эксперимент проводился в течение двух лет (2007–8 годы). Все опыты были проведены не менее чем в 4-х кратной биологической и химической повторностях. Результаты опытов обрабатывались статистически. Данные на рисунках представлены в виде средних значений со стандартными ошибками (n=5)
3. Результаты и их обсуждение
3.1. Влияние абиотических факторов на динамику пигментов фотосинтеза в хвое сосны обыкновенной в зависимости от сезонности
Эволюция фотосинтеза во многом предопределила не только высокий уровень организации растений, но и всех других организмов на Земле. Появление различных модификаций световых и темновых реакций явились одним из ответов на изменяющиеся условия внешней среды и позволили занять растительным организмам различные экологические ниши.
В процессе эволюции у хвойных вечно-зеленых растений в условиях континентального климата выработался ряд адаптивных реакций фотосинтеза. В связи с этим изучение сезонных и возрастных изменений первичных фотосинтетических процессов позволяет исследовать пластические возможности фотосинтетического аппарата и представляет значительный интерес для прикладных эколого-физиологических исследований.
Исследования пигментного состава фотосинтетического аппарата представлены на рис. 1–3; табл. 1. Как видно из рисунков содержание основных и дополнительных пигментов не остается постоянным в течение года. Максимальное их количество отмечается в летние месяцы. Это согласуется с данными , [17 ], согласно которым с переходом в состояние зимнего покоя происходит значительная деградация структуры хлоропластов, приводящая к разрушению пигмент–белковых комплексов (ПБК) и фотоокислению хлорофилла.
По результатам этих исследований и литературным данным можно сказать, что возрастные колебания содержания зеленых пигментов довольно значительны. Для хвои II года характерно более высокое содержание обоих форм хлорофилла и каротиноидов по сравнению с хвоей I года. Первоначально это связано с развитием молодой хвои. Более низкое содержание пигментов в молодой хвое обусловлено с одной стороны тем, что в ней только начинает формироваться фотосинтетический аппарат в летний период ее развития, а с другой стороны частичной деградацией хлоропластов и пигментов в зимний период.
Эту общую закономерность подтверждают данные представленные на рис. 1, которые показывают содержание хлорофилла а в хвое сосны обыкновенной первого и второго года в зависимости от сезонности. Можно констатировать, что с апреля по август месяц происходит повышение содержания хлорофилла а. Именно в этот период начинается активный рост побегов для которых необходим энергетический и пластический материал.
Максимум содержания хлорофилла а приходится на август месяц, когда количество пигмента в хвое 1 и 2-го года выравнивается. Начиная же с сентября месяца происходит постепенное понижение содержания хлорофилла а в хвое как 1-го так и 2-го года. Уменьшение содержания хлорофилла в осенне-зимний период вероятно связано с подготовкой фотосинтетического аппарата хлорофилл-содержащих тканей к зимним условиям при переходе деревьев в состояние покоя. Та же самая зависимость наблюдается и в сезонном изменении содержания хлорофилла в в хвое 1-го и 2-го года (рис.2). Известно, что в зимний период происходит ряд изменений в организации и функционировании фотосинтетического аппарата хвои. Наблюдается пожелтение хвои, обусловленное фотоокислением хлорофиллов [18]. Данный автор считает, что причина зимнего обесцвечивания хлорофилла в том, что низкие температуры при достаточном освещении приводят к диспропорции между световыми и темновыми реакциями фотосинтеза и фотоокислению хлорофилла. Полученные в нашей работе результаты согласуются с данным А. А. Кривошеева, и др. (1991), что сезонные изменения в содержании хлорофилла у сосны обыкновенной обусловлены, с одной стороны, изменениями физиологического состояния деревьев, а с другой – обратимым повреждением хлоропластов хвои в зимний период, поскольку деревья хвойных пород способны переносить низкие отрицательные температуры, сохраняя иглы в течение нескольких лет. При этом следует отметить, что у хвойных вечнозеленых растений было обнаружено наличие зимнего синтеза хлорофилла. В работах показано, что при отрицательной температуре (-20С) происходит включение С14 в молекулу хлорофилла [19]. В связи с этим высказывается предположение о том, что в цепи биосинтеза хлорофилла у хвойных имеется одно или несколько звеньев, образование которых из их предшественников, протекает при температуре не ниже -20 С. [20 ].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
