
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Прямое влияние марганца на синтез хлорофилла связано, прежде всего, с тем, что он входит в состав флавиновых ферментов, участвующих в синтезе порфобиллиногена. Вместе с тем, данный микроэлемент участвует в упрочнении связи хлорофилл–белок, предохраняя пигмент от прежде временного разрушения. Этим можно объяснить повышение содержания марганца в зимние месяцы. При этом учитывается, что изменения активности фотосинтетического аппарата, происходящие в зимний период, обратимы, поскольку деревья хвойных пород приспособлены к переживанию морозов и сохраняют иглы в течение нескольких лет (Барская, 1967).
Как видно из рис. 4 содержание данного микроэлемента больше в хвое II года в зимнее время, чем в хвое I года в этот же период. Затем количество марганца в хвое обоих лет выравнивается. По-видимому, в весенние месяцы энергично синтезируются вещества, идущие на увеличение собственного веса хвои, а в дальнейшем, когда лист достигает нормальных размеров, идет энергичное накопление марганца, а, следовательно, вероятно и активное накопление стрессорных белков. Поэтому хвоя первого года (молодая) чаще подвергается разрушающему действию низких отрицательных температур в отличие от хвои второго года. Полученные результаты совпадают с данными о накоплении марганца растениями в различные месяцы года. [5].
Таким образом, динамика содержания марганца в хвое сосны обыкновенной имеет прямую корреляцию с динамикой отрицательных температур, то есть чем ниже температуры окружающей среды, тем выше концентрация данного микроэлемента в изучаемом объекте.
Представляется интересным сравнение этих данных с результатами, полученными ранее нами, которые свидетельствуют, что подобная закономерность отмечается и в динамике каротиноидов. Динамика зеленых пигментов в те же годы имеет противоположную направленность. Сходная динамика марганца и каротиноидов, по-видимому, не случайна, т. к. известно, что данный микроэлемент повышает содержание каротиноидов. Особое внимание заслуживает тот факт, что марганец способствует увеличению содержания углеводов в иглах хвойных растений. Роль последних в устойчивости клеток к отрицательным температурам общеизвестна.
На основании полученных результатов и литературных данных можно констатировать, что участие марганца в упрочнении связи хлорофилл-белок и предохранение его от разрушения может быть одной из причин устойчивости хвойных к низким температурам и их способности осуществлять фотосинтез даже зимой. Марганец необходим для нормального протекания фотосинтеза, он участвует в процессе на этапе разложения воды и выделения кислорода (фотолиз воды), а также восстановления углекислого газа. Другой причиной устойчивости хвойных растений к низким отрицательным температурам может быть то, что функционально активный марганец в фотосинтетической мембране связан с белками, образуя хелатные соединения. Это дает основание для предположения, что некоторые из этих металлоорганических соединений играют роль в адаптации хвойных растений к низкотемпературному стрессу.
ВЫВОДЫ
1. Содержание зеленых и желтых пигментов не остается постоянным в течение года. Минимальное содержание хлорофиллов отмечается в зимний период, но полного разрушения хлорофиллов в этот период не наблюдается.
2. Одна из причин относительно высокой устойчивости хвойных к низким отрицательным температурам, в частности сосны обыкновенной, может быть обусловлена повышением содержания каротиноидов в зимние месяцы.
3. Повышенное содержание каротиноидов в зимний период обеспечивает повышение температуры хвои на 10–140 С по сравнению с температурой окружающей среды, что способствует осуществлению фотосинтеза в период низких отрицательных температур.
4. Устойчивость хвойных вечнозеленых пород деревьев (сосна обыкновенная) к действию низких отрицательных температур обусловлена не только повышенным содержанием желтых пигментов, но и марганца, содержание которого в зимний период года также наибольшее.
5. Высокое содержание активного марганца, входящего в состав фотосинтетической мембраны и связанного с белками (вероятно стрессовыми белками), по-видимому, способствует адаптации сосны обыкновенной к низким отрицательным температурам.
6. В эксперименте четко прослеживается прямая корреляция между отрицательными температурами и содержанием каротиноидов и марганца. Это указывает на то, что они совместно принимают участие в адаптации хвойных к низким отрицательным температурам.
Литература
1. Полевой растений: Учеб. для биол. спец. вузов - М.: высш. шк., 1989 – 464 с.
2. Очерки об адаптационном синдроме. Пер. с англ. и . – М.: Медгиз, 1960. – С. 254.
3. Болондинский исследования динамики фотосинтеза у сосны обыкновенной// Эколого-физиологические исследования фотосинтеза и водного режима растений в полевых условиях - Иркутск: СиФИБР СО АИ СССР, 1983.
4. Кутюрин воды при фотосинтезе как процесс биологического окисления. - В сб. Механизм дыхания, фотосинтеза и фиксации азота. М.: «Наука», 1967.
5. , Давыдов как микроэлемент в связи с биохимией и свойствами таннидов., Челябинское книжное издательство, 1961.
6. , .Генетические основы радиорезистентности и эволюция. – М..: Энергоатомиздат,1993. – С. 208.
7. Гродзинский растительных систем. – Киев, 1983. – 366 с.
8. Физиология сосны обыкновенной /, , и др.- Новосибирск: Наука. Сибирское отделение, 1990. – 248 с.
9. Физиология древесных растений - М.: Лесная промышленность, 1974. – 420 с
10. Молчанов лес и влага. М.: Наука, 1983. – 139 с.
11. Физиолого-биохимические механизмы роста хвойных. Ответв. редакторы канд. с. - х. наук , канд. биол. наук . Изд-во «Наука» Сибирское отделение. Новосибирск, 1978.
12. Новицкая сосны к экстремальным факторам среды // Физиолого-биохимические основы роста и адаптации сосны на Севере. - Л.: Наука. Ленинградское отделение, 1985. – С. 113–138.
13. , Леванидов двухвалентного марганца перекисью водорода в присутствии цитрат-иона /В сб. Биохимическая роль марганца в растениях. Челябинск, 1967.
14. , Гавриленко и физиология фотосинтеза. Издательство Московского университета 1977.
15. Микроэлементы в жизни растений. – Л.: изд-во «Наука», Ленингр. отд. – 324 с.
16. , , Хандобина практикум по физиологии растений. Фотосинтез. Дыхание. Учебное пособие. М., «Высшая школа», 1975. – 392 с.
17. , Чикина обмен у сосны на Севере. - Л.: Наука. Ленинградское отделение, 1980. – 166 с.
18. Ходасевич аппарат хвойных: Онтогенетический аспект. - Мн: Наука и техника, 1982. – 199 с.
19. Годнев и его роль в природе (конспект лекций по избранным главам физиологии растений). – Минск.: изд-во Белгосуниверситета им. , 1955.
20. Возилова биосинтеза хлорофилла в хвое проростков сосны обыкновенной (автореферат) Томск, 1971.
21. Карнаухов функции каротиноидов. М.: Наука, 1988. –240 с.
22. Дубров и физиологические эффекты действия ультрафиолетовой радиации на высшие растения. М.: Наука, 1968. – 250 с.
23. , Похлебаев -биохимические основы адаптации сосны обыкновенной к экстремальным абиотическим факторам среды обитания. / Экологический вестник Челябинской области. Издательство «СОХА», № 5-6 (16), 2003. – С. 97–109.
24. Крамер, Козловский. Физиология древесных растений / Documents and Settings\Дом.1-ABA9857062AF4, 7 января, 2003.
25. , . Климатогенная реакция деревьев сосны на юге Томской области /Журнал Сибирского Федерального университета: Серия «Биология», т. 1, № 4. – 2008.
ПРИЛОЖЕНИЕ
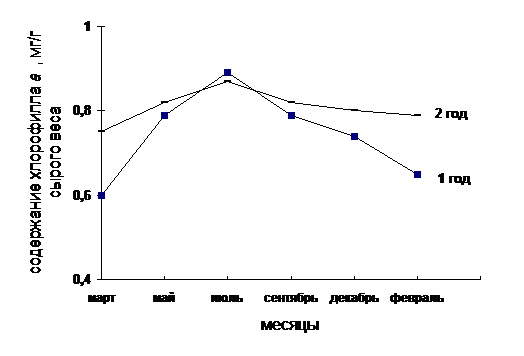
Рис. 1. Содержание хлорофилла а в хвое сосны обыкновенной первого и второго года

Рис. 2. Содержание хлорофилла в в хвое сосны обыкновенной первого и второго года

Рис.3. Содержание каротиноидов в хвое сосны обыкновенной первого и второго года
Таблица 1. Содержанию зеленых и желтых пигментов (мг/г сырого веса) в хвое I и II года сосны обыкновенной в зависимости от сезонности
март | май | июль | сентябрь | декабрь | февраль | |||||||
I г. | II г. | I г. | II г. | I г. | II г. | I г. | II г. | I г. | II г. | I г. | II г. | |
содержание хл.а | 0,6 | 0,75 | 0,79 | 0,82 | 0,89 | 0,87 | 0,79 | 0,82 | 0,74 | 0,8 | 0,65 | 0,79 |
содержание хл в | 0,19 | 0,25 | 0,25 | 0,29 | 0,31 | 0,32 | 0,26 | 0,29 | 0,24 | 0,27 | 0,2 | 0,26 |
содержание каротин-ов | 0,39 | 0,42 | 0,37 | 0,4 | 0,37 | 0,37 | 0,38 | 0,4 | 0,39 | 0,42 | 0,40 | 0,45 |
отношение хл а/хл в | 3,16 | 3 | 3,16 | 2,8 | 2,9 | 2,7 | 3 | 2,8 | 3,1 | 3 | 3,25 | 3,16 |
Таблица 2. Содержание марганца в хвое I года сосны обыкновенной в зависимости от сезонности
№ п/п | месяцы | масса хвои, г | масса золы хвои, г | оптическая плотность, нм | масса Мn, мг/кг а. с.в. |
1 | август | 2,90 | 0,09 | 0,011 | 62,09 |
2 | октябрь | 1,03 | 0,03 | 0,009 | 142,8 |
3 | декабрь | 1,73 | 0,04 | 0,016 | 151,2 |
4 | февраль | 1,21 | 0,04 | 0,011 | 148,9 |
5 | апрель | 1,86 | 0,06 | 0,014 | 123,4 |
6 | май | 3,65 | 0,12 | 0,014 | 64,16 |
Таблица 3. Содержание марганца в хвое II года сосны обыкновенной в зависимости от сезонности
№ п/п | месяцы | масса хвои, г | масса золы хвои, г | оптическая плотность, нм | масса Мn, мг/кг а. с.в. |
1 | август | 2,80 | 0,07 | 0,010 | 66,3 |
2 | октябрь | 1,40 | 0,04 | 0,014 | 164,1 |
3 | декабрь | 1,20 | 0,03 | 0,017 | 232,4 |
4 | февраль | 1,84 | 0,04 | 0,026 | 231,4 |
5 | апрель | 1,56 | 0,05 | 0,014 | 147,3 |
6 | май | 3,28 | 0,11 | 0,014 | 70,1 |
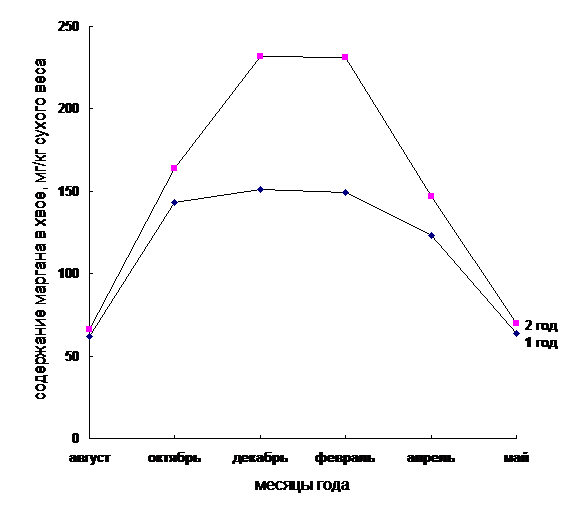
Рис. 4. Содержание марганца в хвое сосны обыкновенной первого и второго года в зависимости от сезонности

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
