

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
1,5 мес | 12 мес | |
контроль |
|
|
|
|
|
Перелом + аМЕМ |
|
|
Перелом + аМЕМ+кмММСК 5%О2 |
|
|
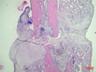
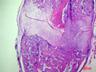

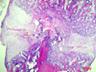
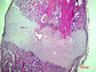
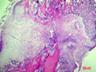
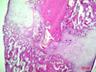

Рис. 6. Костная мозоль на 14 сутки после экспериментального перелома малоберцовой кости крыс. Окраска гематоксилином и эозином. Бар 200 мкм.
1 – костные отломки; 2 – ретикулофиброзная костная ткань; 3 – хрящевая ткань; 4 - неоформленная соединительная ткань
Гистологическое исследование области перелома во всех группах не выявило нарушений процесса заживления кости. Костная мозоль состояла из эндостальной костной мозоли и периостальной костной мозоли, которую покрывала толстая, богатая фибробластами надкостница. Эндостальная костная мозоль состояла из рыхлой неоформленной соединительной ткани, а периостальная костная мозоль включала хрящевую ткань, ретикулофиброзную костную ткань и рыхлую неоформленную соединительную ткань. Во всех исследуемых группах в эндостальной костной мозоли обнаруживались дистальные и/или проксимальные отломки малой берцовой кости, представляющей собой участки пластинчатой костной ткани. Отломки были покрыты остеокластами и окружены новообразованными тканями. Большинство отломков были смещены друг относительно друга. Новообразованная ретикулофиброзная кость располагалась проксимально и дистально относительно хряща. Поверхность трабекул покрывали активные остеобласты.
В результате эксперимента обнаружено увеличение коэффициента утолщения костной мозоли при введении кмММСК, выделенных из костного мозга молодых животных (рис. 7). Причём данное увеличение было отмечено как в группе животных, которым вводили кмММСК молодой крысы, культивированные при 5% кислорода, так и в группе, где были использованы клетки, предварительное культивирование которых осуществлялось в стандартных условиях. Достоверного увеличения коэффициента утолщения при введении клеток, полученных из костного мозга старых животных, выявлено не было.
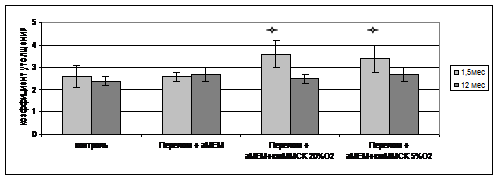
Рис.7. Коэффициент утолщения костных мозолей.
![]() р<0,05 по сравнению с контрольными группами
р<0,05 по сравнению с контрольными группами
При анализе тканевого состава костных мозолей было показано значительное увеличение количества хрящевой ткани после введения кмММСК молодых животных независимо от условий культивирования (табл. 4 и 5). Площадь, занимаемая ретикулофиброзной костной тканью, после введения клеток, культивируемых при 5% кислорода, превышала таковую после введения кмММСК, экспансия которых проходила при 20% кислорода независимо от возраста животного.
Таблица 4. Содержание хрящевой ткани в костной мозоли через 14 суток после экспериментального перелома малой берцовой кости крыс
Группы животных | После введения клеток молодой крысы (1,5 мес) | После введения клеток старой крысы (12 мес) | ||
% от площади мозоли | Площадь ткани в мм2 | % от площади мозоли | Площадь ткани в мм2 | |
контроль | 10,1±2,7 | 2,1±1,1 | 7,1±0,1 | 0,5±0,1 |
Перелом + αМЕМ | 17,2±2,7 | 2,1±0,3 | 5,5±0,7 | 0,4±0,1 |
Перелом + αМЕМ+кмММСК 20%О2 | 24,2±3,0* | 2,6±0,3 | 8,4±1,1** | 0,8±0,1 |
Перелом + αМЕМ+кмММСК 5%О2 | 16,4±1,9 | 1,9±0,2 | 8,6±0,9** | 0,8±0,1 |
* при р<0,05 по сравнению с контрольными группами
** при р<0,05 по сравнению с введением клеток молодой крысы
Таблица 5. Содержание ретикулофиброзной костной ткани в костной мозоли через 14 суток после экспериментального перелома малой берцовой кости крыс
Группы животных | После введения клеток молодой крысы | После введения клеток старой крысы | ||
% от площади мозоли | Площадь ткани в мм2 | % от площади мозоли | Площадь ткани в мм2 | |
контроль | 51,2±4,9 | 5,7±0,8 | 64,5±3,0 | 3,9±0,3 |
Перелом + αМЕМ | 48,3±5,4 | 6,0±0,8 | 58,4±2,1 | 4,2±0,3 |
Перелом + αМЕМ+кмММСК 20%О2 | 48,4±2,6 | 5,4±0,7 | 51,8±2,8 | 3,8±1,2 |
Перелом + αМЕМ+кмММСК 5%О2 | 61,9±2,6 | 7,4±0,6 | 66,9±2,9 | 6,1±0,3 |
Ранее было показано, в том числе и в нашей лаборатории, что культивирование при пониженном содержании кислорода стимулирует пролиферативную активность ММСК первого и последующих пассажей при 5% кислорода (Grayson W. L., et al., 2006; и др., 2007; и др., 2009; Ren H. et al., 2006; Dexheimer V. et al., 2011). Проведенное исследование показало снижение пролиферативной активности в первичной культуре при 5% кислорода во всех возрастных группах, с сохранением тенденции к снижению пролиферативной активности с возрастом. При дальнейшем субкультивировании при 5% кислорода происходило увеличение пролиферативной активности во всех группах, что соответствует ранее полученным результатам (Буравкова и др. 2007). Оценка количества КОЕ-Ф показала, что при постоянном культивировании в условиях 5% кислорода в среде в первичной культуре происходило образование достоверно большего числа колоний по сравнению с 20% кислорода, хотя оценка прироста клеток в фиксированных точках продемонстрировала снижение пролиферативной активности, что связано с разной схемой проведения экспериментов по оценки прироста и выявлению КОЕ-Ф. Оценивая влияние возраста, можно отметить, что независимо от условий культивирования, число КОЕ-Ф кмММСК старых животных значительно ниже, чем среди клеток молодых животных. Tsai C. C. и др. показали, что гипоксия понижает экспрессию E2A-p21 через HIF-1α. Это позволяет сохранять в условиях гипоксии активность теломеразы и длину теломеров ММСК, при этом у клеток был нормальный кариотип и неповрежденный генетический аппарат (Tsai C. C. et al., 2011). Кроме этого, было показано, что высокое содержание HIF-1α может коррелировать со сверхэкспрессией белков-маркеров пролиферации фаз S-G2 клеточного цикла, таких как циклин А и Ki-67, и с активацией клеточной пролиферации ( и др. 2010; Bos R at al., 2004; Gardner L. B. et al., 2003).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
