


Известно, что активность ферментов детоксикации у насекомых [54- 56], равно как чувствительность мишеней для инсектицидов [56, 57], зависит от онтогенетических, возрастных и физиологических особенностей. Так, байгон в 12 раз более токсичен для личинок третьего возраста чувствительной к карбаматам линии М. domestica, чем для имаго [58]. ЛД50 монокротофеса против гусениц S. littoralis четвертого возраста составляет 1447 мкг/г, а против имаго - 19 мкг/г; при добавлении ингибитора активности MM CGA 84708 соответствующие значения составляют 90 и 11 мкг/г. Таким образом, активность микросомальных монооксигеназ является главным фактором устойчивости гусениц S. littoralis к монокротофосу, но оказывает лишь слабое влияние на токсичность инсектицида против имаго [59]. В то же время летальные дозы перметрина и фенвалерата, равно как и КСД их смесей с ППБ против гусениц и имаго устойчивой к пиретроидам полевой популяции колорадского жука, различаются очень слабо [53]. В настоящий момент выявление у устойчивых популяций вредителей сельского хозяйства стадий онтогенеза, чувствительных к химическому контролю, и изучение механизмов устойчивости (в том числе и природной) методами токсикологического анализа является одним из перспективных подходов в борьбе с резистентностью [60].
Половые и возрастные особенности насекомых также могут быть причиной вариабельности значений ЛД50, однако изменения в токсичности инсектицида в известных случаях [35, 58, 61, 62] не всегда коррелируют с изменениями в активности ферментов детоксикации. В любом случае для корректного выполнения токсикологического анализа необходима предварительная селекция биоматериала по возрасту и полу.
Для выявления роли проницаемости покровов в развитии резистентности у насекомых существует три способа. Один из них базируется на использовании различных методов нанесения инсектицида - топикального, инъекции и распыления масляного раствора [63]; если при этом инсектицид детоксицируется гораздо быстрее, чем приходит через кутикулу, то значения ЛД50 для топикального нанесения и инъекции разнятся существенно. Так, при инъекции пиретроидных инсектицидов гусеницам природной популяции S. littoralis, устойчивой к ФОС. их токсичность увеличивалась в 2-3 раза в сравнении с топикальным нанесением; эта популяция обнаружила примерно такой же уровень перекрестной резистентности к пиретроидам [64]. Летальные дозы дильдрина и паратиона для М. domestica в обоих этих методах почти не различались, тогда как токсичность карбарила при инъекции увеличилась более чем в 600 раз. Распыление карбарила с маслом также значительно снижает ЛД50 [63].
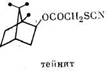 Приведенные данные ставят под сомнение возможность использования методики топикальиого нанесения раствора в ацетоне карбарила и ингибитора ММ для оценки участия этих ферментов в детоксикации инсектицидов, как делают Brattsten и Metcalf [32, 33], Brindley [37] и другие авторы. Ведь если синергист, подобно маслу, способен влиять па проникновение карбарила через покровы насекомого, интерпретация токсикологических данных будет весьма затруднительной и вряд ли корректной. Действительно, Sun и Johnson [65] показали, что для карбаматных инсектицидов степень синергизма с сезамексом на комнатной мухе гораздо выше при топикальном нанесении, нежели при распылении в масле или инъекции. Топикальная токсичность ацетонового раствора SD 8786 (2,3,5-триметилфенил-N-метилкарбамата) значительно увеличивается при добавлении керосина, тогда как КСД для этого соединения с сезамексом при добавлении керосина снижается. Тейнит существенно повышает токсичность карбарила и SD 900 (2,4,5-триметилфенил-N-метилкарбамата) против комнатных мух при топикальном нанесении раствора в ацетоне, но не дает синергизма при распылении в масле [65]; следовательно, тейнит ускоряет проникновение инсектицидов через кутикулу, но не влияет на активность ферментов детоксикации. Эти свойства тейнита позволили Raffa и Priester [66] предложить его для установления роли проницаемости кутикулы в устойчивости насекомых.
Приведенные данные ставят под сомнение возможность использования методики топикальиого нанесения раствора в ацетоне карбарила и ингибитора ММ для оценки участия этих ферментов в детоксикации инсектицидов, как делают Brattsten и Metcalf [32, 33], Brindley [37] и другие авторы. Ведь если синергист, подобно маслу, способен влиять па проникновение карбарила через покровы насекомого, интерпретация токсикологических данных будет весьма затруднительной и вряд ли корректной. Действительно, Sun и Johnson [65] показали, что для карбаматных инсектицидов степень синергизма с сезамексом на комнатной мухе гораздо выше при топикальном нанесении, нежели при распылении в масле или инъекции. Топикальная токсичность ацетонового раствора SD 8786 (2,3,5-триметилфенил-N-метилкарбамата) значительно увеличивается при добавлении керосина, тогда как КСД для этого соединения с сезамексом при добавлении керосина снижается. Тейнит существенно повышает токсичность карбарила и SD 900 (2,4,5-триметилфенил-N-метилкарбамата) против комнатных мух при топикальном нанесении раствора в ацетоне, но не дает синергизма при распылении в масле [65]; следовательно, тейнит ускоряет проникновение инсектицидов через кутикулу, но не влияет на активность ферментов детоксикации. Эти свойства тейнита позволили Raffa и Priester [66] предложить его для установления роли проницаемости кутикулы в устойчивости насекомых.
На определенную роль проницаемости покровов в возникновении у насекомых устойчивости к инсектицидам может указывать также резистентность к оловоорганическим препаратам, например к ацетату трибутилолова [67].
В ряде случаев причиной резистентности насекомых является мутантная АХЭ - мишень действия фосфорорганических и карбаматных инсектицидов, обладающая пониженной чувствительностью к ингибированию этими соединениями. Активность АХЭ и ее чувствительность к ингибиторам определяется только биохимическими методами, поэтому в основе тестирования мутантной АХЭ лежит оценка величины I50 и констант ингибирования in vitro. Однако в ряде случаев возможна токсикологическая оценка чувствительности АХЭ к различным инсектицидам. Так, если (при отсутствии заметного вклада в устойчивость метаболизма и проницаемости инсектицидов) резистентность насекомых к паратиону преодолевается фенитротионом, его этильным аналогом или фентионом, предполагается, что нечувствительность АХЭ обусловливается изменениями в анионной зоне активного центра фермента [68], если же эта резистентность преодолевается метильиым, пропильным и изопропильными аналогами паратиона, анионная зона не включается в изменение чувствительности АХЭ к паратиону [69].
Наиболее полно механизм резистентности за счет мутантной АХЭ был исследован у зеленой рисовой цикадки Nephotettix cincticeps. При сравнении чувствительной и резистентной к карбаматным инсектицидам популяций Yamamoto с соавт. [21] обнаружили существенную разницу в степени ингибирования АХЭ in vitro карбарилом и его N-пропильным аналогом. 1-Нафтил-N-пропилкарбамат оказался в 50 раз менее активным в ингибировании АХЭ из насекомых чувствительной популяции, однако карбарил был в 20 раз менее активным в ингибировании АХЭ насекомых резистентной популяции [21]. Соответственно карбарил был в 25 раз более токсичен на чувствительной популяции, а его пропильный аналог - в 7 раз более токсичен на резистентной в сравнении с чувствительной к карбарилу популяцией. Таким образом, если устойчивость к карбаматным инсектицидам развивается в основном за счет изменений в активном центре АХЭ, сравнительные данные по токсичности карбарила и его аналогов могут дать ответ о наличии или отсутствии резистентности по такому механизму.
Устойчивость насекомых к ДДТ и синтетическим пиретроидам во многих случаях развивается неметаболическим путем, по так называемым kdr - и cynep-kdr-механизмам, связанным с ослаблением взаимодействия инсектицидов с мембраной нервных клеток [70, 71]. Достаточно надежным указанием на участие этого механизма в развитии устойчивости является наличие перекрестной резистентности насекомых к ДДТ и пиретроидам при отсутствии метаболической детоксикации, определяемой применением ингибиторов [72]. Вообще наличие широкого спектра перекрестной устойчивости к пиретроидам дает основание полагать, что эта устойчивость обеспечена kdr-механизмом. Перекрестная же устойчивость с инсектицидами из других химических классов или с иным механизмом токсического действия указывает на наличие метаболических механизмов резистентности или заметный вклад сниженной проницаемости [30].
В простом методе, предложенном Bloomquist и Miller [73], для выявления резистентности насекомых к пиретроидам по kdr-типу используется оценка значений ЭД50 (эффективной дозы пиретроида, вызывающей паралич у 50% особей). Однако и этот способ можно использовать лишь в тех случаях, когда резистентность развивается только по kdr-механизму; альтернативные возможности следует исключать, определяя ЭД50 в присутствии ингибиторов ферментов детоксикации.
Известно, что действие пиретроидов на нервную систему насекомых подчиняется отрицательному температурному коэффициенту, поэтому для определения изменения устойчивости нервной системы можно предложить сравнение токсичности пиретроидных инсектицидов при разных температурах. Действительно, токсичность перметрина на резистентной к перметрину расе S. littoralis при 30° и 10° различается в 12 раз, а на чувствительной - всего в 4 раза [12].
Chang и Plapp [71] показали, что хлордимеформ существенно повышает связывание ДДТ и синтетических пиретроидов с мембраной нервных клеток златоглазки Chrysopa carnea и совки Н. virescens, к тому же хлордимеформ и его аналоги являются синергистами к этим инсектицидам [74]. Возможно, что в будущем хлордимеформ окажется эффективным средством для диагностики kdr-типа резистентности.
Таким образом, нами представлены способы, с помощью которых можно определить, по какому из механизмов у насекомых развивается резистентность к инсектицидам. При этом применение ингибиторов активности ферментов детоксикации для выявления механизмов резистентности является наиболее важной и сложной частью токсикологического анализа.
Значение выявления механизмов резистентности становится особенно очевидным в свете того факта, что устойчивость к одному и тому же инсектициду у различных популяций одного вида может развиваться по разным механизмам [75, 76]. Своевременное определение механизма резистентности позволяет предложить эффективные средства преодоления или задержки развития устойчивости вредителя к инсектициду. Так, Forgash [77] показал, что резистентность колорадского жука к фенвалерату определяется повышением оксидативного метаболизма, после чего применение в полевых условиях смеси фенвалерата с ППБ позволило вести контроль численности этого вредителя [1]. Токсикологический анализ резистентной к ДДТ и пиретроидам популяции моли-пестрянки Phyllonorycter blапсаrdells указал, что резистентность развивается неметаболическим путем; в результате для эффективной ротации были предложены оксамил и метомил [78]. Анализ причин устойчивости различных стадий развития полевой популяции лугового мотылька Loxostege stictlcalis к ФОС и карбаматным инсектицидам позволил выявить наиболее уязвимую для действия токсических агентов имагинальную стадию.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
