


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Агрохимия, 1988, №8, 1988
УДК 632.951: 591.5
ПРИМЕНЕНИЕ ТОКСИКОЛОГИЧЕСКИХ МЕТОДОВ В ИЗУЧЕНИИ МЕХАНИЗМОВ РЕЗИСТЕНТНОСТИ К ИНСЕКТИЦИДАМ У НАСЕКОМЫХ
ЛЕОНОВА И. Н., СЛЫНЬКО Н. М.
Борьба с вредителями сельского хозяйства в настоящее время осуществляется прежде всего с помощью химических средств; возникающая при этом резистентность насекомых к инсектицидам приводит к серьезным экологическим и экономическим проблемам [1].
Основные механизмы защиты насекомых от инсектицидов включают в себя изменение проницаемости покровов, увеличение активности ферментных систем, принимающих участие в детоксикации инсектицидов, и изменение чувствительности мишени для токсических веществ [2]. Известно, что один и тот же механизм может обеспечивать развитие устойчивости к химическим соединениям разных классов и способствовать возникновению перекрестной резистентности к ним.
Резистентность к инсектицидам может иметь моно- и многофакторный характер, когда один или большее число механизмов отвечают за ее возникновение к одной и той же группе инсектицидов. Эффективность защитных возможностей насекомых определяет порядок, в котором они селектируются под влиянием отбора. При этом сочетание различных механизмов устойчивости приводит к значительному повышению, уровня резистентности к инсектицидам.
Существует несколько способов, с помощью которых можно существенно затормозить развитие резистентности и, соответственно, повысить действенность применяемых химических средств борьбы с вредителями сельского хозяйства. Во-первых, создание новых эффективных и высокоселективных инсектицидов, с каждым годом требующее все больших затрат времени и средств [3], во-вторых, чередование применения инсектицидов, или ротация [4], и, наконец, в-третьих, использование синергистов, которые в смеси с инсектицидами увеличивают их токсичность [5]. Применение в сельском хозяйстве последних двух методов требует своевременной диагностики резистентности популяции вредителей к инсектицидам, а также точного представления о механизмах ее возникновения.
Для выявления механизмов резистентности насекомых к инсектицидам применяются два подхода. Один из них включает определение активности основных систем детоксикации in vitro, изучение их изоферментного состава, чувствительности ацетилхолинэстеразы (АХЭ) к ингибиторам [6]. Другой, являющийся предметом настоящего обзора, включает в себя токсикологические методы: оценку токсического действия различных инсектицидов на разных стадиях развития насекомых; проведение анализа с использованием соединений, которые, проникая в организм насекомых, специфически тормозят активность ферментов детоксикации и в сочетании с инсектицидом увеличивают его токсическое действие. По усилению токсичности под действием таких специфических ингибиторов можно судить об участии тех или иных ферментов в метаболизме инсектицидов.
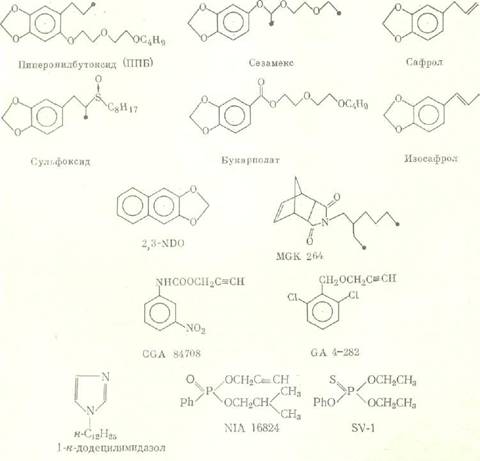
Микросомальные монооксигеназы (ММ) являются одной из наиболее важных ферментных систем, определяющих развитие резистентности насекомых к инсектицидам. Вследствие того что ММ катализируют протекание множества реакций и характеризуются широкой субстратной специфичностью, они принимают участие в метаболизме карбаматов, пиретроидов, ряда хлорорганических и фосфорорганических инсектицидов и обусловливают возникновение перекрестной резистентности [7]. Для доказательства роли ММ в ингибиторном анализе используются соединения из различных химических классов.
Химическое строение некоторых ингибиторов активности ММ:
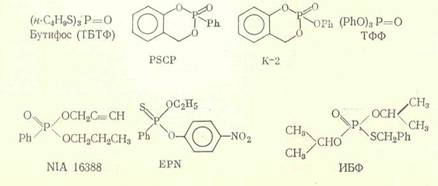
В развитии резистентности насекомых к инсектицидам немаловажную роль играют неспецифические эстеразы, осуществляющие гидролиз многих фосфорорганических соединений и пиретроидов [8]. Для эстераз также доказано наличие разных изоформ, характеризующихся специфичностью к гидролизу определенных субстратов. Однако среди употребляемых в токсикологическом анализе ингибиторов эстераз практически нет соединений, влияющих именно на одну из реакций.
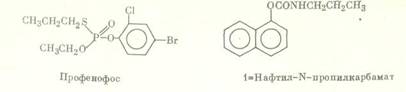
Химическое строение некоторых ингибиторов активности эстераз:
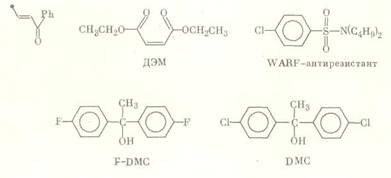
Довольно ограниченный набор химических соединений используется для выявления роли глутатион S-трансфераз и ДДТ-дегидрохлориназ, которые, как показано, также обеспечивают развитие резистентности к некоторым ФОС и ДДТ [9, 10].
Химическое строение ингибиторов активности глутатион S-трансфераз и ДДТ-дегидрохлориназы:
Первый этап токсикологического анализа - определение токсичности инсектицида для данной популяции вредителя и, если это возможно, сравнение с данными, полученными на чувствительной линии. Затем в зависимости от химического класса инсектицида проводят испытания для смесей инсектицида с подходящим синергистом: так, карбаматные инсектициды обычно испытывают только с ингибиторами монооксигеназной активности, пиретроиды - с ингибиторами ММ и карбоксилэстераз, а ФОС - с ингибиторами ММ, неспецифических эстераз и глутатион S-трансфераз. Для выяснения механизма устойчивости вредителя к инсектициду важен как факт наличия синергизма или антагонизма, так и их отсутствия, Данные по отсутствию синергизма, однако, не дают оснований сделать вывод о том, что исследуемая ферментная система не принимает участия в метаболизме инсектицида, и могут быть обусловлены неудачным выбором ингибитора метаболизма инсектицида, снижением скорости прохождения инсектицида через покровы, метаболизмом синергиста у данного вида вредителя и т. п.
Важна для корректного проведения ингибиторного анализа специфичность ингибиторов активности ферментов детоксикации, которая может зависеть от различных факторов, например, от видовых особенностей или количества нанесенного на насекомых вещества. Так, ППБ, селективно, как правило, ингибирующий активность микросомальных монооксигеназ, в высоких концентрациях способен влиять на активность неспецифических эстераз. n-Додецилимидазол является ингибитором активности ММ различных видов насекомых и млекопитающих [11], однако в эксперименте было показано, что in vitro он в концентрации 5-10-7 М в 5 раз снижает активность карбоксилэстераз, выделенных из гусениц резистентной к пиретроидам популяции хлопковой листовой египетской совки Spodoptera littoralis, и не влияет на активность эстераз из гусениц резистентной карадрины Spodoptera exagua [12]. Большинство исследователей считают бутифос специфичным ингибитором эстераз [13], однако у гусениц южной абмарной огневки Plodia interpunctella он подавляет и монооксигеназную активность [14]. Ингибитор эстераз NIA 16388 синергизирует биоресметрин и перметрин на резистентных к пиретроидам линиях комнатной мухи Musca domestica [15]; однако на том основании, что совместное синергистическое действие ППБ и NIA 16388 неаддитивно, авторы делают вывод, что последнее соединение ингибирует в данном случае оба фермента. Для доказательства участия глутатион 5-трансфераз в резистентности насекомых к инсектицидам многие авторы в качестве ингибитора активности этого фермента используют диэтилмалеат (ДЭМ). Однако в ряде работ показано, что ДЭМ оказывает заметное влияние на активность монооксигеназ, так что достоверные данные об активности трансфераз с его использованием можно получить лишь в том случае, если предварительные испытания с ингибитором активности ММ (ППБ, сезамекс) не обнаружили синергизма [16].
Кроме того, ингибиторы ферментов детоксикации могут оказывать влияние на неметаболические процессы распределения инсектицидов в организме членистоногих. ППБ тормозит проникновение аллетрина через кутикулу [17], тогда как бутифос, напротив, ускоряет проницаемость трихлорфона [18]. В литературе имеются данные, что ППБ, являясь эффективным синергистом перметрина на М. domestica, не оказывал никакого влияния на метаболические процессы [19]. Ингибитор ДДТ-дегидрохлориназы DMC способен влиять на взаимодействие ДДТ и пиретроидов с нервным рецептором [20], а ингибитор неспецифических эстераз 1-нафтил-N-пропилкарбамат оказался токсичным для вредителей риса, резистентность которых к фосфорорганическим и карбаматным инсектицидам возникла в результате мутации ацетилхолинэстераз [21].
Выбор оптимального синергиста для данного инсектицида на конкретном виде вредителя проводится, как правило, эмпирически и осложняется ингибиторной специфичностью применяемых веществ, обусловленной существованием изоформ ферментов детоксикации инсектицидов [22]. Например, ингибиторы активности монооксигеназ ППБ, SV-1 и NIA 16824 заметно синергизировали токсичность цис-циперметрина на личинках хрущака Tribolium castaneum, однако только NIA 16824 синергизировал транс-перметрин [23]. Тот же NIA 16824 был весьма эффективным синергистом цис - и транс-тетраметрина на М. domestica, в то время как на других видах насекомых его влияние, равно как и прочих ингибиторов ММ, было малозначимым [24]. На резистентной к пиретроидам линии М. domestica GA 4-282 проявляет больший синергистический эффект с карбаматами, а ППБ - с перметрином и ресметрином [25].
Синергистическое действие ИБФ, бутифоса и т. п. обычно связывают с их способностью ингибировать гидролиз соединений, содержащих в своей структуре карбоксильные или карбоксиамидные фрагменты. Несмотря на то, что роль гидролаз в метаболизме метилпаратиона и других ФОС, не содержащих карбоксильные группы, ставится под сомнение [26], бутифос и EPN синергизируют метилпаратион на резистентной популяции табачной совки Heliothis virescens [27]. Аналогичный результат получен при изучении высокорезистентной к темефосу линии комара Culex quinquefasciatus, устойчивость которой в значительной степени подавляется применением бутифоса и ИБФ, тогда как трифенилфосфат проявляет лишь небольшой эффект [28]. Данные по синергизму метилпаратиона и темефоса указывают на то, что диапазон действия применяемых ингибиторов не ограничен подавлением активности карбоксилэстераз и карбоксиамидаз, поскольку эти инсектициды соответствующих групп в своей структуре не содержат.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
