

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
- роль ферментов метаболизма цитокининов (синтеза, распада и модификации), в частности аденозинкиназ, в передаче гормонального сигнала и жизнедеятельности растительной клетки;
- цитокинин-связывающие белки растений: их изучение, выделение и функции;
- гормон-связывающие глютатион-трансферазы и их роль в растении.
Вторая глава
Содержит описание объектов и методов исследования:
Объектами исследования были выбраны растения с расшифрованными последовательностями генома: Arabidopsis thaliana L., экотип Colambia (3 и 6-недельные розетки листьев); Oryza sativa L., сорт «Юпитер» (листья 4-х недельных растений). Также в качестве объектов использовались Hordeum vulgare L., сорт «Луч» (закончившие рост 9-10-дневные листья первого яруса); Vigna radiata L. (3-х дневные этиолированные проростки).
Применялось несколько схем выделения белков. Для выделения цитокинин-связывающих белков сначала проводили гель-фильтрацию экстракта из цитозоли или ядер растений на сефадексе G-50 и гидрофобную хроматографию на фенил-сефарозе, затем раствор белков наносили на специально синтезированные аффинные смолы зеатинрибозид-сефарозу или глютатион-сефарозу. Цитокинин-связывающие белки вытесняли с зеатинрибозид-сефарозы неспецифически щелочью или специфически: гормонами и лигандами. в низкой концентрации (0,5-5мМ), чтобы предупредить неспецифическое вытеснение, связанное с увеличением ионной силы раствора. Очищенные таким образом белки тестировались методом твердофазного иммуноферментного анализа на нитроцеллюлозной мембране или полистироловом планшете с помощью антиидиотипических антител к транс-зеатину, антиидиотипических антител к ИУК, антител на пептид N-конца АСБ65 (CASTKRVLV). Антиидиотипические антитела являются вторичными антителами к антителам на гормон и представляют собой антитела к гормон-связывающему участку белка.
Для поиска новых цитокинин-связывающих белков Arabidopsis thaliana L. и Oryza sativa L. проводилось специфическое вытеснение белков этих растений с зеатинрибозид-сефарозы цитокинином транс-зеатином в низкой концентрации (1мМ), электрофорез в денатурирующих условиях полученной цитокинин-связывающей фракции белков. Идентификация белков по результатам MALDI масс-спектрометрии белковых полос осуществлялась с использованием поисковой системы Mascot (www. ) в доступной базе данных NCBInr для Arabidopsis thaliana L. и Oryza sativa L.
Для определения степени сродства белков аденозинкиназ Hordeum vulgare L. к различным производным цитокининов и другим молекулам, проводилась вытеснение этими лигандами в равной концентрации (0,5мМ) белков с аффинной смолы зеатинрибозид-сефарозы.
Третья глава
Содержит результаты проведенных исследований и их обсуждение.
3.1. Взаимодействие ЦСБ67 с антителами к консервативной последовательности гистидинкиназ.
К настоящему времени описано семейство мембранных гибридных гистидинкиназ Arabidopsis thaliana L. (AHK2, AHK3, AHK4), причем одна из них (CRE1/WOL/AHK4) является мембранным рецептором цитокинина. Поликлональные антитела на консервативный пептид гистидинкиназ не взаимодействовали с выделенным ЦСБ67 (см. рис.2, трек 5). Кроме того, в отличие от гистидинкиназ AHK, ЦСБ67 не является мембранным белком. Таким образом, ЦСБ67 не относится к белкам класса гистидинкиназ.
3.2. Изучение свойств цитокинин-связывающего белка ЦСБ67 и ауксин-связывающего белка АСБ65 Джонса.
ЦСБ67 из 10-дневных листьев Hordeum vulgare L., полученный путем неспецифического вытеснения щелочью с зеатинрибозид-сефарозы, взаимодействовал с антителами на пептид АСБ65, а также с антиидиотипическими антителами к ИУК (рис.2). Также был выделен АСБ65 из 3-дневных этиолированных проростков Vigna radiata L. по методу Джонса, с помощью очистки на Q-сефарозе, который взаимодействовал на иммуноблоте с антиидиотипическими антителами к транс-зеатину (рис.1). Таким образом, каждый из белков АСБ65 и ЦСБ67 содержит три идентичные иммунодетерминанты, так как взаимодействует с тремя типами используемых антител: с антиидиотипическими антителами к транс-зеатину, с антиидиотипическими антителами к ИУК и с антителами на пептид АСБ65 (CASTKRVLV). При этом исключена вероятность того, что общие иммунодетерминанты сходны с бычьим сывороточным альбумином, к которому пришивались гормоны и пептид для иммунизации, так как ЦСБ67 не взаимодействует с антителами к пептиду из 19 аминокислот консервативного домена гистидинкиназ (рис.2), которые также были получены иммунизацией кроликов конъюгатом пептида с бычьим сывороточным альбумином.
Так как АСБ65 задерживается глютатион-сефарозой и N-концевая последовательность АСБ65 высоко гомологична некоторым из растительных глютатион-трансфераз, было сделано предположение, что ЦСБ67 также будет связываться с глютатион-сефарозой. С помощью глютатион-сефарозы была получена фракция белков из ядер 10-дневных листьев Hordeum vulgare L., связавшаяся с этой смолой и специфически вытесняемая со смолы восстановленным глютатионом в низкой концентрации (5мМ). Эта фракция использовалась для выделения цитокинин-связывающих белков с помощью аффинной хроматографии на зеатинрибозид-сефарозе с неспецифическим вытеснением щелочью. В результате был получен белок, который определен как ЦСБ67 (рис.3), поскольку он взаимодействовал с антиидиотипическими антителами к транс-зеатину, а также усиливал синтез РНК in vitro до 300% в совокупности с транс-зеатином в транскрипционной системе, содержащей хроматин и связанную с ним РНК-полимеразу I из листьев Hordeum vulgare L. Следовательно, ЦСБ67 является глютатион-связывающим белком.
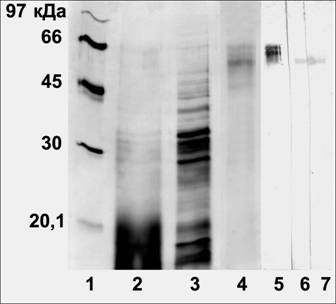
Рис. 1. 1-4: очистка АСБ65 Vigna radiata L., электрофорез в ДДС-Na-ПААГ, окраска серебром; 5-7: иммуноблоты очищенного АСБ65. 1- маркеры; 2 - фракция, осажденная сульфатом аммония; 3 - фракция после очистки на фенил-сефарозе; 4 - фракция после очистки на Q-сефарозе; 5 - антитела к пептиду АСБ65; 6 - антиидиотипические антитела к ИУК; 7 - антиидиотипические антитела к транс-зеатину.
Было проведено определение глютатион-трансферазной активности очищенного ядерного и цитозольного ЦСБ67 в реакции с субстратом CDNB in vitro. Ферментативной активности обнаружено не было (табл.1). Для АСБ65 также известно, что он не проявляет глютатион-трансферазной активности с CDNB, однако показывает незначительную активность (120% по сравнению с контролем) с субстратом металохлором. Таким образом, нельзя исключать, что АСБ65 и ЦСБ67 обладают глютатион-трансферазной активностью с другими субстратами. Также вероятно, что они теряют способность проявлять глютатион-трансферазную активность в результате модификации белков при их выделении.
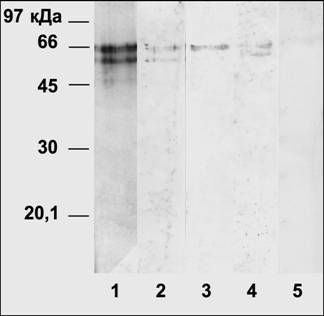
Рис. 2. ЦСБ67 Hordeum vulgare L. 1: электрофорез в ДДС-Na-ПААГ, окраска Кумасси; 2-5: иммуноблоты очищенного ЦСБ67. 1 - очищенный ЦСБ67; 2 - антиидиотипические антитела к транс-зеатину; 3 - антитела к пептиду АСБ65; 4 - антиидиотипические антитела к ИУК; 5 – антитела к консервативной последовательности гистидинкиназ.

Рис. 3. Очистка ЦСБ67 из ядер Hordeum vulgare L. с помощью глютатион-сефарозы. 1-3: электрофорез в ДДС-Na-ПААГ, окраска серебром, 4: иммуноблот. 1 – белок ядер Hordeum vulgare L., 2 – фракция после очистки на глютатион-сефарозе, 3 – фракция после очистки на зеатинрибозид-сефарозе; 4 – иммуноблот выделенного белка с антиидиотипическими антителами к транс-зеатину.
В результате проведенных исследований было сделано предположение, что АСБ65 и ЦСБ67 принадлежат к одному классу растительных белков, т. к. обладают одинаковыми структурными и функциональными характеристиками и, таким образом, они являются аналогичными белками, выделенными из разных видов растений, Hordeum vulgare L. и Vigna radiata L.
Табл. 1. Глютатион-трансферазная-активность ЦСБ67, выделенного из ядер Hordeum vulgare L. и из цитозоли Hordeum vulgare L., с субстратом CDNB (данные усреднены по 10 повторностям).
Контроль | ЦСБ67 из ядер | ЦСБ67 из цитозоли | |
Оптическая плотность, 340 нм | 0,180 | 0,189 | 0,182 |
Значения в %, относительно контроля | 100 | 105 | 101 |
На основании того, что ЦСБ67 и АСБ65 имеют одинаковые свойства, также было сделано предположение, что ЦСБ67 связывает не только гормон цитокинин, но и ауксин. Для выяснения того, связываются цитокинин и ауксин одним участком ЦСБ67 или разными, проведилось вытеснение антиидиотипических антител к транс-зеатину с помощью транс-зеатина и ИУК в системе конкурентного твердофазного иммуноферментного анализа на микропланшетах. При увеличении концентрации транс-зеатина уже до 10-8М, он практически полностью вытеснял антиидиотипические антитела к транс-зеатину (рис.4). Тогда как ИУК во всем исследуемом диапазоне концентраций 10-9-10-5М не вытесняла антиидиотипические антитела к транс-зеатину. Следовательно, если ЦСБ67 связывает гормоны транс-зеатин и ИУК, то связывание осуществляется разными участками этого белка.
Антитела на пептид N-конца белка АСБ65 CASTKRVLV не вытеснялись транс-зеатином и ИУК во всем исследуемом диапазоне концентраций гормонов (10-9-10-5М) в системе конкурентного твердофазного ИФА (рис.5). Это говорит о том, что участок ЦСБ67, гомологичный пептиду CASTKRVLV и взаимодействующий с антителами на пептид АСБ65, не участвует в связывании ауксина и цитокинина.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
