

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
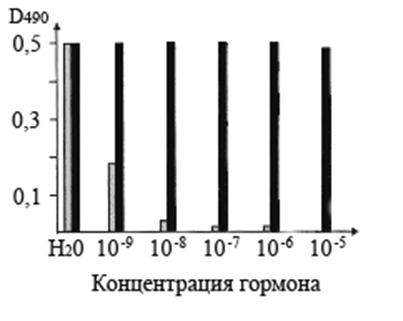
Рис. 4. Вытеснение антиидиотипических антител к транс-зеатину, связанных с ЦСБ67 Hordeum vulgare L., транс-зеатином (серые столбики) и ИУК (черные столбики).
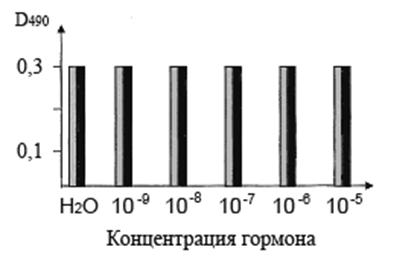
Рис. 5. Вытеснение антител к пептиду АСБ65, связанных с ЦСБ67 Hordeum vulgare L., транс-зеатином (серые столбики) и ИУК (черные столбики).
ЦСБ67 и АСБ65 связывают восстановленный глютатион, гомологичны растительным глютатион-трансферазам класса phi, при этом не обнаружена глютатион-трансферазная активность ЦСБ67 и АСБ65 с субстратом CDNB in vitro. Однако надо отметить, что в отличие от большинства глютатион-трансфераз, для которых характерна молекулярная масса 24-26кДа, АСБ65 и ЦСБ67 имеют молекулярную массу 65-67кДа.
Согласно данным последних лет, глютатион-трансферазы являются не просто участниками в детоксикации ксенобиотиков, а полифункциональными белками с самыми разнообразными функциями. Предполагаются следующие функции связывания гормонов ауксинов и цитокининов растительными глютатион-трансферазами: 1) участие в передаче гормонального сигнала в растении путем гормон-зависимого связывания с протеинкиназами, возможность чего показано на глютатион-трансферазах животных; 2) участие в транспорте гормонов; 3) защита гормонов от метаболизации неспецифическими оксидазами путем его обратимого связывания; 4) регуляция уровня свободного гормона внутри клетки путем его обратимого связывания; 5) модуляция ферментативной активности глютатион-трансфераз гормоном in vivo.
3.3. Изучение активации транскрипции белком ЦСБ67 in vitro.
Ранее было показано, что ЦСБ67 в совокупности с транс-зеатином усиливает синтез РНК in vitro в 2-3 раза в транскрипционной системе, содержащей хроматин и связанную с ним РНК-полимеразу I из листьев Hordeum vulgare L. Согласно результатам данной работы, ЦСБ67 из ядер 6-недельных розеток листьев Arabidopsis thaliana L. усиливает синтез РНК in vitro более чем в 3 раза в совокупности с транс-зеатином в транскрипционной системе, содержащей хроматин и связанную с ним РНК-полимеразу I из листьев Hordeum vulgare L. (табл.2). Таким образом, цитокинин-связывающие белки ЦСБ67, выделенные из разных системных групп растений (Hordeum vulgare L. – Класс Однодольные; Arabidopsis thaliana L.– Класс Двудольные) являются консервативными, а также консервативны транскрипционные системы, выделенные из этих объектов.
Выделенный с помощью глютатион-сефарозы и зеатинрибозид-сефарозы из ядер 10-дневных листьев Hordeum vulgare L., ЦСБ67 усиливал синтез РНК in vitro до 300% в совокупности с транс-зеатином в транскрипционной системе, содержащей хроматин и связанную с ним РНК-полимеразу I из листьев Hordeum vulgare L.
АСБ65, выделенный с помощью преципитации сульфатом аммония, фенил-сефарозы, Q-сефарозы, не усиливал синтез РНК in vitro в совокупности с транс-зеатином в транскрипционной системе, содержащей хроматин и связанную с ним РНК-полимеразу I из листьев Hordeum vulgare L. Предполагается, что полученный таким образом белок не был активен, так как в схеме его очистки применялось осаждение сульфатом аммония.
После предобработки ЦСБ67 в эквимолярном количестве реактивом Эллмана, блокирующим свободные SH-группы и добавлении его в систему транскрипции in vitro, усиления транскрипции не наблюдалось (табл.2). Сделано предположение, что для проявления активности ЦСБ67, необходимы его свободные SH-группы.
Таблица 2. Влияние ЦСБ67 из ядер 6-недельных розеток листьев Arabidopsis thaliana L. на синтез РНК in vitro в транскрипционной системе, содержащей хроматин и связанную с ним РНК-полимеразу I из листьев Hordeum vulgare L.
ЦСБ67, нг | транс-зеатин, 10-7М | Включение [Н3] УМФ | |
Имп/мин на 10 мкг ДНК | % | ||
- | - | 26784 | 10 |
- | + | 25844 | 97 |
20 | - | 30773 | 115 |
20 | + | 97891 | 319 |
40 | + | 79338 | 297 |
20 (предобработан реактивом Эллмана) | - | 19980±486 | 75 |
20 (предобработан реактивом Эллмана) | + | 18962±531 | 71 |
3.4. Поиск и идентификация новых цитокинин-связывающих белков Arabidopsis thaliana L. и Oryza sativa L.
Была проведена очистка белков, специфически связывающих транс-зеатин, из растений с расшифрованной последовательностью генома: 4-недельных листьев Oryza sativa L. и 3-недельных розеток листьев Arabidopsis thaliana L. В отличие от схемы очистки для ЦСБ67, где белки с аффинной смолы зеатинрибозид-сефарозы неспецифически вытеснялись щелочью, в данном случае проводилась специфическое вытеснение — раствором транс-зеатина в низкой концентрации (1мМ). Таким образом, можно однозначно заключить, что полученные белки являются цитокинин-связывающими in vitro.
С помощью масс спектрометрии белковых полос цитокинин-связывающей фракции, для Arabidopsis thaliana L. в полосах 150, 80, 45, 42кДа (рис.6) была идентифицирована смесь двух изоформ белков аденозинкиназ: АДК1 (ген At3g09820) и АДК2 (At5g03300). При этом для АДК1 и АДК2 в этих полосах счёт составлял от 82 до 182 (при достоверно значимом счёте более 60), за исключением полосы 80кДа, где счет был 42 для АДК2 и 40 для АДК1, и другие белки для этой полосы также не были достоверно определены. Белки АДК1 и АДК2 во всех полосах, где были идентифицированы, присутствовали в смеси. Об этом говорит тот факт, что среди определенных пептидов, были как общие для родственных белков АДК1 и АДК2, так и специфические только для АДК1 или АДК2. Так, например, в полосе для Arabidopsis thaliana L. 42кДа было определено 10 пептидов, общих для АДК1 и АДК2; 7 пептидов, специфических только для АДК2; 6 пептидов, специфических только для АДК1. Таким образом, в полосах 150, 80, 45, 42кДа белки АДК1 и АДК2 присутствуют в смеси, т. е. оба этих белка имеют сходные структурные свойства и являются цитокинин-связывающими in vitro.
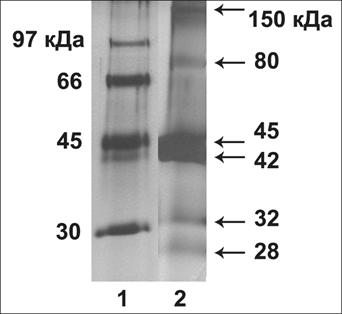
Рис. 6. Выделение цитокинин-связывающих белков из 3-недельных розеток листьев Arabidopsis thaliana L.. Электрофорез в ДДС-Na-ПААГ, окраска серебром. 1 – маркеры; 2 – фракция, вытесненная с зеатинрибозид-сефарозы транс-зеатином 1мМ.
Для Oryza sativa L. был выявлен и достоверно определен белок аденозинкиназа с молекулярной массой 42кДа (Os02g0625500) (рис.7). Тогда как для Arabidopsis thaliana L. с помощью аффинной смолы зеатинрибозид-сефарозы очищались белки АДК1 и АДК2 в смеси, то для Oryza sativa L. с помощью метода фингерпринта была определена только одна из двух аденозинкиназ, чьи гены присутствуют в геноме. Можно предположить, что ген второй аденозинкиназы Oryza sativa L. (Os04g0518000) не экспрессируется в растении, или его продукт не выделятся с помощью используемой для очистки схемы, так как его структурные и функциональные свойства значительно отличаются от продукта гена Os02g0625500.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
