

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
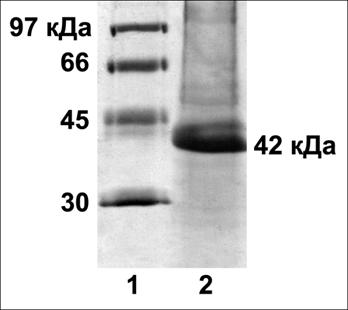
Рис. 7. Выделение цитокинин-связывающих белков из 30-дневных листьев Oryza sativa L. Электрофорез в ДДС-Na-ПААГ, окраска серебром. 1 – маркеры; 2 – фракция, вытесненная с зеатинрибозид-сефарозы транс-зеатином 1мМ.
Ранее было показано, что аденозинкиназы связываются с зеатин-сефарозой и вытесняются с нее транс-зеатином, фосфорилируют аденозин, дигидрозеатин, изопентениладенин, зеатинрибозид in vitro. Роль связывания ферментами аденозинкиназами цитокининов может заключаться: в преобразовании рибозидов и оснований цитокининов в соответствующие нуклеотиды, что происходит, например, при добавлении экзогенных цитокининов; в поддерживании оптимального уровня активных форм цитокининов в клетке; в обеспечении регуляции метаболизма цитокининов, связанного с клеточным циклом растительных клеток. Белки аденозинкиназы являются белками «домашнего хозяйства», их гены экспрессируются конститутивно, белки присутствуют в клетках в значительном количестве и являются абсолютно необходимыми для обеспечения жизнедеятельности клетки.
Белок полосы 32кДа Arabidopsis thaliana L. предположительно определен как протеинкиназа НТ1 (ген At1g62400).
Белок полосы 28кДа Arabidopsis thaliana L. достоверно определен как тиаминаза (ген At3g16990). Ген At3g16990 соответствует гибридному эукариотическому белку ThiD-TenA, который содержит на С-конце фрагмент, схожий с бактериальным активатором транскрипции (TenA), и на N-конце фрагмент, родственный HMP-P киназам, вовлеченным в биосинтез тиамина/витамина В1 (ThiD). Тиаминазы растений не изучены, известна третичная структура тиаминазы, в результате определения которой исследователями была обнаружена плотность в структуре белка, которая вставлена между двумя ароматическими кольцами и имеет размер и форму пурина или индола. Было сделано предположение, что плотность может образовывать молекула растительного гормона – цитокинина или ауксина, что согласуется с полученными нами данными о связывании тиаминазой гормона транс-зеатина in vitro. Физиологический смысл этого связывания остается неизвестным.
3.5. Вытеснение белка аденозинкиназы Hordeum vulgare L. с зеатинрибозид-сефарозы различными производными цитокининов и гомологичными им молекулами.
Фракция гидрофобных белков из 10-дневных листьев Hordeum vulgare L. наносилась зеатинрибозид-сефарозу. Специфическое вытеснение аденозинкиназы проводилась в промывочном растворе с добавлением лиганда в концентрации 0,5-1мМ.
На рис.8 и рис.9 видно, что транс-зеатин и АТФ вытесняли сходное количество белка аденозинкиназы с зеатинрибозид-сефарозы. Может существовать два объяснения этому факту: АТФ и транс-зеатин связываются в одном месте аденозинкиназы, и добавление свободного АТФ в вытесняющий раствор вытесняет транс-зеатин из места связывания, освобождая белок. Также возможно, что связывание АТФ и транс-зеатина осуществляется в разных местах, и вытеснение с помощью АТФ происходит в результате изменения конформации белка при связывании АТФ с АТФ-связывающим участком. Количество вытесненного со смолы белка прямо пропорционально количеству добавленного в вытесняющий раствор лиганда, причем степень точности достаточно высока (см. рис.8, 9). Таким образом, количество белка аденозинкиназы, вытесненного со смолы лигандом, пропорционально двум факторам: количеству добавленного в вытесняющий раствор лиганда, а также степени сродства лиганда к белку.
Было проведено вытеснение белков аденозинкиназ Hordeum vulgare L. с зеатинрибозид-сефарозы с помощью различных производных цитокининов и гомологичных им молекул в концентрации 0,5мМ (см. рис.10, 11). Количество белка аденозинкиназы, вытесненного транс-зеатином было принято за 100%. Данные обработаны с помощью программы 1DScan EX.
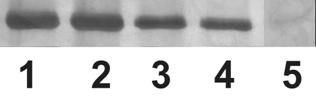
Рисунок 8. Вытеснение аденозинкиназы Hordeum vulgare L. с зеатинрибозид-сефарозы добавлением транс-зеатина и АТФ в разных концентрациях: 1 - транс-зеатин 1мМ; 2 - АТФ 1мМ; 3 - транс-зеатин 0,5мМ; 4 - АТФ 0,5мМ; 5 - контроль (промывочный раствор). Электрофорез в ДДС-Na-ПААГ, окраска серебром.
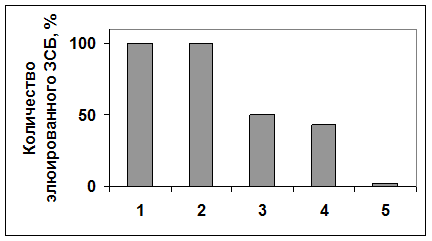
Рисунок 9. Относительное количество вытесненного белка аденозинкиназы Hordeum vulgare L. (по данным рис.9). Количество белка, вытесненного транс-зеатином 1мМ, принято за 100%. 1 - транс-зеатин 1мМ; 2 - АТФ 1мМ; 3 - транс-зеатин 0,5мМ; 4 - АТФ 0,5мМ; 5 - контроль.
Согласно полученным результатам, аденозинкиназа Hordeum vulgare L. имеет сходную степень сродства к транс-зеатину и зеатинрибозиду, а также к аденину и аденозину, т. е. появление рибозной группы в молекуле пурина не влияет на степень сродства белка с гормоном, что согласуется с функцией аденозинкиназы - фосфорилированием рибозидов с использованием АТФ в качестве донора фосфорила, и свободных оснований производных аденина с использованием фосфорибозила. Таким образом, наличие или отсутствие рибозного остатка в молекуле пурина не изменяет значимо степень сродства белка к лиганду.
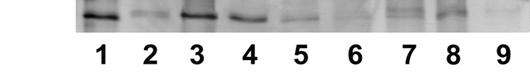
Рисунок 10. Вытеснение аденозинкиназы Hordeum vulgare L., связанной с зеатинрибозид-сефарозой, различными лигандами (0,5мМ): 1 - зеатинрибозид; 2 - аденозин; 3 - АТФ; 4 - транс-зеатин; 5 - аденин; 6 - 6-БАП; 7 - ГТФ; 8 - УТФ; 9 - контроль (промывочный раствор). Электрофорез в ДДС-Na-ПААГ, окраска серебром.
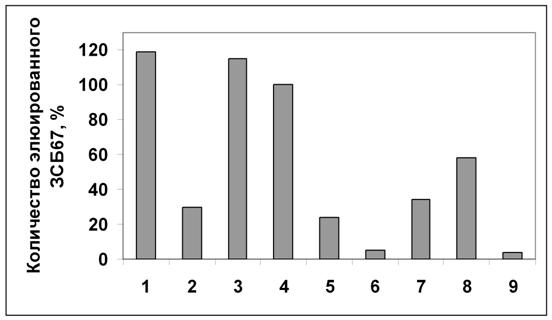
Рисунок 11. Относительное количество вытесненного белка аденозинкиназы Hordeum vulgare L. (по данным рис.10). Количество белка, вытесненного транс-зеатином 0,5 мМ, принято за 100%. 1 - зеатинрибозид; 2 - аденозин; 3 - АТФ; 4 - транс-зеатин; 5 - аденин; 6 - 6-БАП; 7 - ГТФ; 8 - УТФ; 9 - контроль.
Аденин и аденозин, которые считаются классическим субстратом аденозинкиназы, снимают примерно в 4 раза меньше белка, чем транс-зеатин и его производные. Известно, что субстратом аденозинкиназы in vitro являются аденозин и некоторые производные цитокининов (дигидрозеатин, изопентениладенин, зеатинрибозид), но активность фермента с использованием в качестве субстратов цитокининов, ниже, чем с использованием аденозина. Согласно полученным нами результатам, аденозинкиназа связывает транс-зеатин с большей степенью сродства, чем аденозин.
Синтетический цитокинин 6-БАП не вытесняет белок аденозинкиназу с зеатинрибозид-сефарозы. Следовательно, в результате замены боковой группы транс-зеатина на бензольное кольцо, нарушается связывание белка с лигандом, при этом отсутствие радикала в этом положении (как у аденина) уменьшает сродство белка к лиганду примерно в 4 раза, но не полностью. Это согласуется с тем, что искусственный цитокинин 6-БАП вызывает физиологический эффект в растении (вероятно, путем взаимодействия с цитокининовыми рецепторами), однако в отличие от натуральных цитокининов, 6-БАП является высоко стабильным в растении. Можно предположить, что такая особенность объясняется тем, что 6-БАП не используется в качестве субстрата ферментами метаболизма цитокининов, в частности, не фосфорилируется аденозинкиназой.
ГТФ вытесняет со смолы примерно в 3 раза и УТФ в 2 раза меньше аденозинкиназы, чем АТФ. Т. е., сродство белка к другим нуклеотидам значительно ниже по сравнению со сродством к АТФ.
Выводы
1. Установлено, что ЦСБ67 Hordeum vulgare L. не имеет структурного сродства с мембранным рецептором цитокининов гистидинкиназой CRE1/WOL/AHK4 Arabidopsis thaliana L..
2. Установлено, что цитокинин-связывающий белок ЦСБ67 Hordeum vulgare L. и ауксин-связывающий белок Джонса Vigna radiata L. имеют сходные структурные и функциональные свойства, а именно: молекулярную массу, три общие иммунодетерминанты, связывание глютатиона, отсутствие глютатион-трансферазной активности.
3. Обнаружено сродство и возможная идентичность ЦСБ67 Hordeum vulgare L. и АСБ65 Vigna radiata L., а также способность ЦСБ67 связывать гормон ауксин.
4. Установлена функциональная идентичность ЦСБ67 и транскрипционных систем, выделенных из растений, относящихся к разным таксономическим группам: Классу Однодольных (Hordeum vulgare L.) и Классу Двудольных (Arabidopsis thaliana L.).
5. Определена первичная структура цитокинин-связывающих белков Arabidopsis thaliana L.: АДК1 (At3g09820), АДК2 (At5g03300), тиаминазы (At3g16990) и предположительно протеинкиназы НТ1 (At1g62400); а также цитокинин-связывающего белка Oryza sativa L.: АДК (Os02g0625500).
6. Определена степень сродства цитокинин-связывающего белка аденозинкиназы Hordeum vulgare L. к природному цитокинину: зеатинрибозиду, синтетическому цитокинину: 6-БАП, а также к производным аденина и нуклеотидам: аденину, аденозину, АТФ, ГТФ; УТФ, по сравнению со сродством к природному транс-зеатину.
Публикации по материалам диссертационной работы
1. , Новейшие достижения в изучении механизма действия фитогормонов // Биохимия. 2004. Т.69. №3. C. 293-311.
2. , , Диффузия органического катиона в клеточных стенках корня // Биохимия. 2003. Т. 68. №7. C. 926-940.
3. Selivankina S. Yu., Karavaiko N. N., Maslova G. G., Zubkova N. K., Prokoptseva O. S., Smith A. R., Hall M. A., Kulaeva O. N. (2004) Cytokinin-binding protein from Arabidopsis thaliana leaves participating in transcription regulation // Plant Growth Regulation. V. 43. pp. 15-26.
4. , , Кулаева характеристика ядерного цитокинин-связывающего белка (67КДА) // Сборник трудов V Съезда Общества физиологов растений. C. 470. Пенза, 15-21 сент. 2003г.
5. , , Кондаков зеатин-связывающих белков и идентификация их генов // Сборник трудов Международной научной конференции «Ломоносов-2008». C. 292. Москва, 7-11 апр. 2008.
6. , , Кондаков сродства белков аденозинкиназ ячменя к производным растительных гормонов цитокининов и гомологичным им молекулам // Тез. докл. III Всероссийской конференции — школы «Высокореакционные интермедиаты химических реакций и биологических процессов». — Астрахань – Москва. C. 20.
7. , , , , (2007) Метод определения индивидуальной чувствительности к пищевым продуктам и коррекции патологических состояний мелких домашних и сельскохозяйственных животных // Методические указания для ветеринарных врачей. Министерство сельского хозяйства РФ, Санкт-Петербург.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
