

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ISBN 978-9939-50-172-7
Баскевич М.И., Потапов С.Г., Хляп Л.А., Малыгин В.М., Опарин М. Л.., 2011. Генетическое и таксономическог разнообразие одноцветных мышовок (RODENTIA, DIPODOIDEA, SICISTA) Кавказа в свете кариологических и молекулярно-генетических данных. Исторические реконструкции и современное состояние популяций // Матер. Межд. Конф. «Биологическое разнообразие и проблемы охраны фауны Кавказа» (Ереван, 26 сент-30 сент. 2011 г.). 72-77
Матер. Межд. Конф. «Биологическое разнообразие и проблемы охраны фауны Кавказа» (Ереван, 26 сент-30 сент. 2011 г.). Ереван: Изд-во «Асогик»
ГЕНЕТИЧЕСКОЕ И ТАКСОНОМИЧЕСКОЕ РАЗНООБРАЗИЕ ОДНОЦВЕТНЫХ МЫШОВОК (RODENTIA, DIPODOIDEA, SICISTA) КАВКАЗА В СВЕТЕ КАРИОЛОГИЧЕСКИХ И МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИХ ДАННЫХ. ИСТОРИЧЕСКИЕ РЕКОНСТРУКЦИИ И СОВРЕМЕННОЕ СОСТОЯНИЕ ПОПУЛЯЦИЙ
1, 1, 1, 2, 1
1Институт проблем экологии и эволюции им. РАН, 119071 Москва, Ленинский пр. 33, e-mail: mbaskevich@mail.ru, 2МГУ им. М. В, Ломоносова, 119992, Москва, Воробьевы горы 1/12, e-mail: *****@***ru
Прослушать
На латинице
GENETIC AND TAXONOMIC DIVERSITY OF UNSTRIPPED BIRCH MICE OF THE CAUCASUS (RODENTIA, DIPODOIDEA, SICISTA) IN THE LIGHT OF CHROMOSOMAL AND MOLECULAR-GENETIC DATA. HISTORICAL RECONSTRUCTION AND CURRENT STATUS OF POPULATIONS
Baskevich M. I.1, Potapov S. G.1, Khlyap LA1, Malygin VM 2, Oparin M. L1
1 Severtsov Institute of Ecology and Evolution RAS, 119071 Moscow, Leninsky Prospekt 33, e-mail: *****@***ru, 2 Moscow State University by M. V, Lomonosov Moscow Vorobyovi Gori 1/12, e-mail: *****@***ru
Прослушать
На латинице
За почти полтора века изучения представления о таксономическом разнообразии одноцветных мышовок Кавказа неоднократно изменялись: их рассматривали в составе S. concolor Buchner (Satunin, 1896 - цит. по Огнев, 1948), признавали в качестве самостоятельного вида S. caucasica Vinogr. (Vinogradov, 1925), возвращались к первоначальной точке зрения (Кузякин, 1963 - цит. по Соколов и др., 1981), а ныне трактуют в виде группы географически замещающих видов-двойников. В соответствии с современными представлениями группа одноцветных мышовок Кавказа объединяет 4 кариологически дискретных вида-двойника и 6 кариоморф: S. caucasica Vinogr. (2n=32, NF=48, 2n=32, NF=46), S. kluchorica Sokol. et al. (2n=24, NF=48), S. kazbegica Sokol. et al. (2n=42, NF=52, 2n=40, NF=50), S. armenica Sokol.& Baskevich (2n=36, NF=52) (Соколов и др.,1981, 1986, Соколов, Баскевич, 1988, 1992, Баскевич, Малыгин, 2009). Хромосомные подходы не только легли в основу серии таксономических ревизий группы, они внесли весомый вклад в изучение закономерностей формирования ее таксономического и генетического разнообразия. Кариологическое изучение новых находок одноцветных мышовок Кавказа представляет безусловный интерес, а степень дифференциации между кариологически дискретными видами-двойниками и различными кариоморфами мышовок, не совпадающая с их морфологическими отличиями, а также филогенетические связи в группе и ее положение в системе рода Sicista нуждаются в оценке с помощью не только хромосомных, но и новых подходов. К их числу могут быть отнесены молекулярно-генетические методы исследования, имевшие до сих пор весьма ограниченное применение в изучении таксономии и филогении рода (Баскевич и др., 2004).
ЦЕЛЬ настоящего исследования - сравнительные хромосомные (G-, C-, AgNOR-banding) и молекулярно-генетические (сиквенс-анализ фрагмента /1099 п. н/ мт гена cyt b) исследования 4-х видов Sicista, в т. ч. 3-х видов-двойников, относящихся к группе одноцветных мышовок Кавказа: S. kluchorica, S. kazbegica. S. caucasica.
МАТЕРИАЛ И МЕТОДЫ
В работе были использованы 9 экз. мышовок, перечень которых дан в таблице 1
Все использованные в работе выборки мышовок были кариологически датированы. Препараты митотических хромосом получали в соответствии со стандартной методикой воздушно-высушенных препаратов. Для изученных форм выполнена в соответствии с общепринятыми методами дифференциальная окраска хромосом (G-, C-, AgNOR-окраски).
Геномную ДНК выделяли из фиксированных в 96% спирте образцов печени с использованием стандартного метода, включающего лизис ткани раствором, содержащим 100 мл мМ Трис-НСl (pH=8), 10 мМ ЭДТА, 100 мМ NaCl, 1% додецилсульфат натрия и протеиназу К (Sigma, USA) в концентрации 0,2 мк/мл при 56°С в течение 12-16 ч. С последующей депротеинизацией смесью фенола и хлороформа. Качество и количество ДНК оценивали с использованием метода электрофореза в 1% агарозном геле и детекции ДНК в УФ-свете после окрашивания геля бромистым этидием. Ген цитохрома b амплифицировали в полимеразной цепной реакции (ПЦР) в виде двух перекрывающихся фрагментов ДНК с использованием праймеров L14724 и H15915 (Irwin et al., 1991 – цит. по Fan et al., 2009). Амплификацию проводили в течение 40 циклов с температурным профилем: 94ºC – 1 мин, 46ºС – 1 мин 30 с, 72°С – 1 мин 30 с. Продукты амплификации очищали с помощью ультрафильтрации на колонках Microcon (Amicon, USA). Секвенирование гена цитохрома b проводили с помощью автоматического секвенатора АВI Prism TM 377 с использованием набора для геномного секвенирования BigDye Terminator v. 3.1 Cycle Sequencing Kit (Appllied Biosystems) и праймеров L14724, H15915 (Irwin, 1991– цит. по Fan et al., 2009), а также L23; H23 (Fan et al., 2009). Использовали фрагмент гена цитохрома b размером 1099 п. н..
Для выравнивания и анализа полученных последовательностей применен пакет программ Sequence Navigator (Applied Biosystems), а для филогенетического анализа – пакет филогенетических программ MEGA 3.1. Генетические дистанции (p-дистанции) между индивидуальными последовательностями ДНК рассчитывали, исходя из количества нуклеотидных замен на позицию при по-парных сравнениях.
Филогенетические деревья, учитывающие характер гаплотипической изменчивости фрагмента митохондриального гена cyt b, построены с применением UPGMA и NJ алгоритмов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Использование разработанных нами ранее хромосомных маркеров (Соколов и др., 1981, 1986; Соколов, Баскевич, 1988, 1992) позволило подтвердить принадлежность мышовок из нового пункта обследования в верховьях р. Сказдон (Цейское ущ., Северо-Осетинский заповедник) в Северной Осетии к 40-хромосомной S. kazbegica (NF=50), ранее зарегистрированной нами в том же ущелье, но в долине р. Цейдон (табл. 1) (Соколов, Баскевич, 1992). Эти результаты подтверждают приуроченность 40-хромосомной S. kazbegica к терскому варианту поясности. Обнаружение клухорской мышовки (2n=24, NF=48) в ущ. Адыл-Су (Национальный Парк «Приэльбрусье») в Кабардино-Балкарии, подтверждает литературные данные (Дзуев, 1988 – цит. по Баскевич и др., 2004) и уточняет восточную границу распространения этого вида-двойника одноцветных мышовок Кавказа, ареал которого на северных склонах Большого Кавказа простирается по хромосомным данным к западу до Клухорского перевала в Карачаево-Черкессии (эльбрусский вариант поясности) (Баскевич и др., 2004). Как известно, западнее на Большом Кавказе обитает 32-хромосомный вид-двойник S. caucasica (Соколов и др., 1981; Баскевич и др., 2004; Баскевич, Малыгин. 2009), чья кариологически типированная находка из верховий р. Мзымта в Сочинском р-не Краснодарского края использована в настоящем сообщении. К сожалению, в данной работе отсутствует сравнительный материал по 36-хромосомному виду-двойнику, S. armenica, пока известному лишь из одного пункта на Малом Кавказе в северо-западной Армении (окрестности пос. Анкаван Разданского р-на) (Соколов, Баскевич, 1988). Полученные нами хромосомные результаты согласуются с сформированными ранее представлениями о видовом разнообразии и распространении видов-двойников и кариоморф одноцветных мышовок Кавказа (Баскевич и др., 2004; Баскевич, Малыгин, 2009).
Детали дифференциальной окраски хромосом у изученных нами экз. видов-двойников одноцветных мышовок Кавказа дополняют полученные на других выборках хромосомные результаты (Баскевич и др., 2004). Показано, что наиболее обособлены среди сравниваемых форм 40-хромосомная S. kazbegica (древняя) и 24-хромосомная S. kluchorica (самая молодая в группе): их G-окрашенные кариотипы различаются восемью не-робертсоновскими транслокациями, а также двумя перицентрическими инверсиями. Подтверждены резкие различия в характере С- и AgNOR-окраски хромосом между S. kazbegica, с одной стороны, и S. kluchorica и S. caucasica, с другой, свидетельствующие в пользу подразделенности одноцветных мышовок с Большого Кавказа на западную и восточную группы.
Результаты молекулярно-генетических (cyt b) исследований Sicista приведены ниже. Перечень использованных образцов Sicista и их краткие хромосомные характеристики представлены в таблице 1.
Таблица 1. Список использованных в молекулярно-генетических исследованиях кариологически типированных образцов видов-двойников одноцветных ышовок Кавказа (1-7) и использованных в качестве внешней группы образцов S. subtilis (8, 9)
№ п/п | Наиме- нование образца | Видовая принадлежность | 2n/NF* | Локалитет |
1 | 10-39 | S. kluchorica | 24/44 | Кабардино-Балкария, ущ. Адыл-Су |
2 | 10-40 | S. kluchorica | 24/44 | - « - |
3 | 10-67 | S. kluchorica | 24/44 | -« - |
4 | 10-114 | S. kluchorica | 24/44 | - « - |
5 | 10-82 | S. kazbegica | 40/52 | Cеверная Осетия, Цейское ущ., верховья р. Сказдон |
6 | 10-89 | S. kazbegica | 40/52 | - « - |
7 | 7-49 | S. caucasica | 32/46 | Краснодарский край, Сочинский р-н, верховья р. Мзымта |
8 | 6-65 | S. subtilis | 23/43 | Саратовская обл., Воскресенский р-н, окрестности пос. Славянка |
9 | 7-24 | S. subtilis | 24/41 | - « - |
* 2n - диплоидное число хромосом, NF - число плеч хромосом
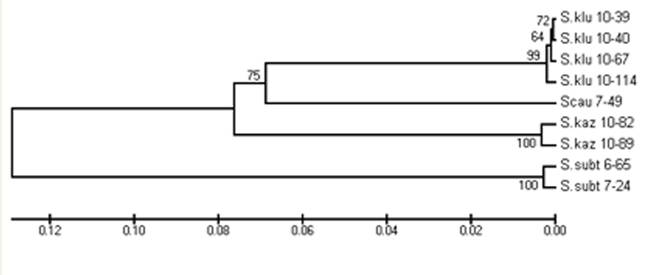
Анализ филогенетических древ, построенных на основе сопоставления значений генетических дистанций между сравниваемыми видами Sicista с помощью UPGMA и NJ алгоритмов, указывает на значительную дифференциацию между одноцветными мышовками Кавказа и исследованным представителем группы subtilis, S. subtilis. Следующий уровень кластеризации соответствует подразделенности на виды внутри группы одноцветных мышовок Кавказа. При этом среди трех сравниваемых видов группы S. kazbegica наиболее обособлена от двух других более близких видов (S. kluchorica, S. caucasica) (табл. 2, рис. 1).
Таблица 2. Генетические дистанции (d) между четырьмя видами Sicista, рассчитанные на основе использования фрагмента (1099 п. н.) митохондриального гена cyt b с помощью К2Р (Кимура 2 параметрического алгоритма)
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
1 | 0.0009 | 0.0012 | 0.0020 | 0.3756 | 0.4532 | 1.1444 | 0.6336 | 0.6210 | |
2 | 0.0009 | 0.0015 | 0.0023 | 0.3755 | 0.3745 | 0.3168 | 0.6339 | 0.6213 | |
3 | 0.0018 | 0.0027 | 0.0015 | 0.3757 | 0.3749 | 0.3393 | 0.6331 | 0.6205 | |
4 | 0.0046 | 0.0055 | 0.0027 | 0.3756 | 0.3750 | 0.3392 | 0.6332 | 0.6206 | |
5 | 0.1517 | 0.1528 | 0.1516 | 0.1504 | 0.0026 | 0.4530 | 0.7266 | 0.7236 | |
6 | 0.1441 | 0.1452 | 0.1440 | 0.1428 | 0.0064 | 0.4430 | 0.7017 | 0.6987 | |
7 | 0.1356 | 0.1367 | 0.1379 | 0.1394 | 0.1733 | 0.1722 | 0.7479 | 0.7448 | |
8 | 0.2456 | 0.2470 | 0.2457 | 0.2457 | 0.2798 | 0.2726 | 0.2760 | 0.0024 | |
9 | 0.2424 | 0.2438 | 0.2425 | 0.2425 | 0.2795 | 0.2723 | 0.2757 | 0.0055 |
Примечание. Под диагональю значения генетических дистанций (d), над диагональю – значения соответствующих стандартных ошибок (S. E.). Образцы S. kluchorica обозначены цифрами:1 (10-39), 2 (10-40), 3 (10-67), 4 (10-114); S. kazbegica - 5 (10-82), 6 (10-89); S. caucasica – 7 (7-49); S. subtilis – 8 (6-65), 9 (7-24). В скобках показаны номера образцов Sicista, соответствующие таковым на рисунке 1.
Рис. 1. Филогенетические деревья, построенные с применением NJ (вверху) и UPGMA (внизу) алгоритмов, учитывающие характер гаплотипической изменчивости фрагмента (1099 п. н.) гена cyt b у 4-х видов Sicista: 3-х представителей группы одноцветных мышовок Кавказа (S. caucasica, S. kluchorica, S. kazbegica) и изученных в качестве внешней группы представителей S. subtilis. В узлах ветвления показано значение буттстреп поддержки для 1000 нуклеотидных замен. Наименование образцов соответствует таковому в таблице 1. Шкала генетических дистанций показана в средней части рисунка
Полученные нами приоритетные молекулярные данные согласуются с результатами сравнительной кариологии. Так, согласно представленным на исследованных выборках молекулярно-генетическим данным, прослеживается поддержка видовой дифференциации в группе, а также подразделенность видов-двойников одноцветных мышовок с Большого Кавказа на западную, и восточную группы. Этот результат, вероятно, связан с событиями плейстоценовой истории и может рассматриваться в рамках аллопатрической модели формообразования и становления ареалов видов-двойников в группе одноцветных мышовок Кавказа. Использование в качестве внешней (базальной) группы образцов S. subtilis, известной из отложений позднего плиоцена Восточной Европы (Топачевский, 1965) может служить подтверждением выше упомянутой гипотезы. В то же время, принимая во внимание, жесткую связь в распространении видов-двойников и кариоморф одноцветных мышовок Кавказа с вариантами поясности, можно высказать предположение о взаимосвязи эволюции этой известной с плиоцена (Верещагин, 1959) группы Кавказских грызунов с формированием высотно-поясной структуры Кавказа. Следует также подчеркнуть, что современное распространение одноцветных мышовок Кавказа связано исключительно с ненарушенными ландшафтами высокогорий и среднегорий Кавказа (оптимум ареалов в субальпийском поясе), сохранившимися лишь на охраняемых территориях (изученные нами находки могут служить подтверждением). Длительные (на протяжении веков) негативные антропогенные воздействия являются дополнительным фактором, определяющим мозаичный характер ареалов видов-двойников и их демовую структуру. Однако ни на хромосомном ни на молекулярно-генетическом уровне различия между малыми популяциями видов-двойников пока не найдены.
NJ

UPGMA
Авторы выражают глубокую благодарность за содействие в организации работы , , за помощь в сборе материала , ,
Исследование поддержано РФФИ (09-04-00464а), Программой «Биоразнообразие (2.6.2) и грантом (2009-1.1-141-063-021) ФЦП «Научные и научно-педагогические кадры инновационной России»
ЛИТЕРАТУРА - 11
Баскевич В. М. 2009. Хромосомные подходы в изучении закономерностей формирования генетического и таксономического разнообразия грызунов Кавказа на примере мышовок, Sicista (Rodentia, Dipodoidea) фауны Кавказского региона//В кн: Горные экосостемы и их компоненты. М.: Изд-во КМК. Ч. 2: 204-210.
Баскевич Н. М., , 2004. Диагностика, распространение и эволюция одноцветных мышовок Кавказа (Rodentia, Dipodoidea, Sicista) // Зоол. Журн. 83 (2):.220-233.
1959. Млекопитающие Кавказа. М.-Л. Изд-во АН СССР. 703 с.
1948. Звери СССР и прилежащих стран. М.-Л. Изд-во АН СССР. 6: 1-706.
, 1988. Новый вид одноцветных мышовок (Rodentia, Dipodoidea) с Малого Кавказа// Зоол. Журн. 67 (2): 300-304.
, 1992. Новая хромосомная форма одноцветных мышовок из Северной Осетии (Rodentia, Dipodoidea, Sicista) // Зоол. журн. 71 (8): 94-103.
, , 1981. Ревизия одноцветных мышовок Кавказа: виды-двойники Sicista caucasica Vinigradov, 1925 и S. kluchorica sp. n. (Rodentia, Dipodidae) // Зоол. журн. 60 (9): 1386-1393.
, , 1986. Sicista kazbegica sp. n. (Rodentia, Dipodidae) из бассейна верхнего течения реки Терек//Зоол. журн. 65 (6): 949-952.
, 1965. Насекомоядные и грызуны ногайской позднеплиоценовой фауны. Киев: Наукова Думка. 163 с.
Fan Z., Sh. Liu, Y. Liu, Bo Zeng, X. Zhang, C. Guo, B. Yue, 2009. Molecular phylogeny and taxonomic reconsiderationof the subfamily Zapodinae (Rodentia: Dipodidae), with an emphasis of Chinese species // Molecular Phylogenetics and Evolution 51: 447-453.
Vinogradov B. S. 1925. On the structure of the external genitalia in Dipodidae and Zapodidae (Rodentia) as a classificatory character // Proc. Zool. Soc. London. 95: 582-584.
SUMMARY
Представлены новые данные по хромосомной (рутинная, G-, C0, AgNOR-окраска хромосом) и молекулярно-генетической (cytb) изменчивости 3-х видов-двойников группы одноцветных мышовок Кавказа (S. kluchorica, S. kazbegica, S. caucasica) и используемого в качестве внешней группы S. subtilis. Дана оценка вклада двух примененных нами генетических подходов в изучение таксономического и генетического разнообразия данной группы Sicista. Исторические реконструкции в группе и современное состояние популяций видов-двойников одноцветных мышовок Кавказа обсуждаются в свете полученных хромосомных и молекулярно-генетических данных.
