

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Использование различных моделей и методологии моделирования необходимо для того, чтобы:
· понять, как устроен конкретный объект, каковы его структура, основные свойства, законы развития и взаимодействия с окружающим миром;
· научиться управлять объектом (процессом) и определить наилучшие способы управления при заданных целях и критериях;
· прогнозировать прямые и косвенные последствия реализации разных способов и форм воздействия на объект.
Хорошо построенная модель, как правило, дает новые знания об объекте-оригинале. Это, безусловно, очень важное свойство, стимулирующее развитие методов моделирования.
Более подробно о моделях и моделировании можно узнать из литературы, например [10]. Далее приведем несколько простейших моделей из экологии и безопасности жизнедеятельности.
9.2. Динамика популяций
В современной экологии часто возникает вопрос: как определить численность той или иной популяции через определенное время? Ответ на него не только представляет теоретический, интерес, но и имеет большое практическое значение. Действительно, не зная этого, нельзя правильно планировать эксплуатацию различных возобновляемых природных ресурсов – промысловых рыб, охотничьих угодий и т. п. Может ли в решении этого вопроса помочь математика? Оказывается, да. Рассмотрим здесь некоторые простейшие модели, на которых проиллюстрируем подход к данному вопросу.
Пусть некоторая популяция имеет в момент времени t0 биомассу x0. Предположим, что в каждый момент времени скорость увеличения биомассы пропорциональна уже имеющейся биомассе, а возникающие явления конкуренции за источниками питания и самоотравления снижают биомассу пропорционально квадрату наличной биомассы. Если обозначить биомассу в момент времени t через х(t), а изменение ее за время ![]() t через
t через ![]() х, то можно записать следующее приближенное равенство:
х, то можно записать следующее приближенное равенство:
![]() х≈(kх-αх2)
х≈(kх-αх2) ![]() t, (9.1)
t, (9.1)
где α и k – положительные постоянные (параметры).
В дифференциальной форме это соотношение имеет вид:
![]() . (9.2)
. (9.2)
Оно и представляет собой математическую модель процесса изменения биомассы популяций. В экологической литературе уравнение (9.2) часто называют логистическим.
Если теперь поставить вопрос о том, какова же будет биомасса в момент времени Т, то на него можно ответить экспериментально – дождаться этого момента и определить биомассу непосредственным измерением (вообще говоря, такое измерение может быть физически неосуществимым).
Другой путь – воспользоваться математической моделью, решая задачу Коши для уравнения (9.2) с начальным условием (9.3):
x(t0)=x0. (9.3)
Разделяя в уравнении (9.2) переменные, получим уравнение в дифференциалах
![]() . (9.4)
. (9.4)
Для дальнейшего удобно ввести новую переменную
z=αх, (9.5)
тогда (9.4) можно переписать в виде
![]()
![]() (9.6)
(9.6)
Возвращаясь к исходному уравнению (9.2), заметим, что если x0=![]() (т. е. z0=k), то задача Коши имеет решение x(t)
(т. е. z0=k), то задача Коши имеет решение x(t)![]() x0 (рис. 9.1). Если x0 <
x0 (рис. 9.1). Если x0 <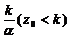 , то уравнение (9.6) интегрируется следующим образом
, то уравнение (9.6) интегрируется следующим образом
ln z – ln(k-z)=ln z0- ln (k-z0)+k(t-t0),
откуда
![]() , (9.7)
, (9.7)
значит,
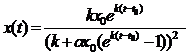 , t > 0 (9.8)
, t > 0 (9.8)
Если x0 > ![]() , то аналогично предыдущему случаю снова получаем формулу (9.8). Дифференцируя (9.8) по t, имеем
, то аналогично предыдущему случаю снова получаем формулу (9.8). Дифференцируя (9.8) по t, имеем
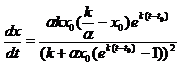 , (9.9)
, (9.9)
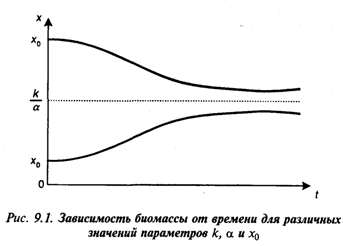
откуда вытекает, что при x0 < ![]() график функции х(t) монотонно возрастает, а при x0>
график функции х(t) монотонно возрастает, а при x0>![]() – монотонно убывает, причем оба графика имеют горизонтальную асимптоту х=
– монотонно убывает, причем оба графика имеют горизонтальную асимптоту х=![]() (рис. 9.1). Мы не приводим здесь элементарную, но громоздкую формулу второй производной d2x/dt2, показывающую, что верхний и нижний графики имеют по одной точке перегиба.
(рис. 9.1). Мы не приводим здесь элементарную, но громоздкую формулу второй производной d2x/dt2, показывающую, что верхний и нижний графики имеют по одной точке перегиба.
Мы рассмотрели весьма упрощенную ситуацию, так как предполагали, что популяция не взаимодействует ни с какими другими популяциями, учет же этого обстоятельства, конечно, значительно усложняет модель.
Рассмотрим одну из таких моделей. Будем обозначать биомассы двух популяций через х и у соответственно. Предположим, что обе популяции потребляют один и тот же корм, количество которого ограничено, и из-за этого находятся в конкурентной борьбе друг с другом.
Французский математик В. Вольтерра в 1926 г. показал, что при таком предположении динамика популяций достаточно хорошо описывается следующей системой дифференциальных уравнений:
![]() , (9.10)
, (9.10)
где 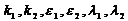 – определенные положительные числа.
– определенные положительные числа.
Первые члены правых частей системы (9.10) характеризуют скорость роста популяций при отсутствии ограничивающих факторов. Вторые члены учитывают те изменения в скоростях, которые вызываются ограниченностью корма.
Задавая различные значения параметров, с помощью системы (9.10) можно описать взаимодействие двух популяций, одна из которых – хищник, а другая – жертва [36]. В литературе [47] более подробно описаны математические аспекты исследования системы (9.10).
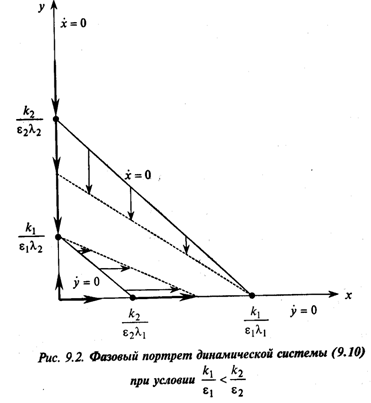
Прежде чем исследовать, как будет вести себя система (9.10), заметим, что в любой момент времени t ее состояние полностью описывается значениями х и у: каждому состоянию системы соответствует некоторая точка (х, у) на плоскости хОу, называемой «фазовой плоскостью». Каждой точке фазовой плоскости можно поставить в соответствие вектор (стрелку на рис. 9.2) с координатами, которые являются правыми частями системы, указывающий направление движения в этой точке. Проведя из начальной точки линии, касательные этим векторам, получим траектории, по которым будет происходить движение системы, т. е. решения задачи Коши для системы (9.10) с начальными условиями
x(t0)=x0, y(t0)=y0, (х0,у0)Î х0у. (9.11)
Чтобы составить представление о траекториях движения системы, построим линии, на которых х=0 (здесь векторы параллельны оси Оу) и у = 0 (здесь векторы параллельны оси Ох). Для краткости обозначим производную ![]() – через х, а
– через х, а ![]() – через у. Имеем
– через у. Имеем
х=0, когда ![]() ,
,
у=0, когда ![]() ,
,
т. е. х = 0 на двух прямых в фазовой плоскости:
х=0 и ![]() =
=![]() ,
,
а у=0 также на двух прямых:
у=0 и ![]() =
=![]() (рис. 9.2, 9.3).
(рис. 9.2, 9.3).
По этим рисункам можно сделать следующие выводы. В обоих случаях имеем три стационарные точки, в которых одновременно х=0 и у=0, а именно: (0,0), (0, ![]() ) и (0,
) и (0, ![]() ), которые по известной классификации являются узлами. При этом, если
), которые по известной классификации являются узлами. При этом, если ![]() >
> ![]() (рис. 9.2), то устойчивым является только узел (
(рис. 9.2), то устойчивым является только узел (![]() , 0), а если
, 0), а если ![]() <
< ![]() (рис. 9.3), то узел (0,
(рис. 9.3), то узел (0, ![]() ). Таким образом, если
). Таким образом, если ![]() >
> ![]() , то вторая популяция вымирает, y(t) → 0, t →
, то вторая популяция вымирает, y(t) → 0, t → ![]() , а первая стабилизируется, x(t) →
, а первая стабилизируется, x(t) →![]() , t →
, t →![]() . Если же
. Если же ![]() <
< ![]() , то имеем обратную картину: первая популяция вымирает, x(t) → 0, t→
, то имеем обратную картину: первая популяция вымирает, x(t) → 0, t→![]() , а вторая стабилизируется, x(t) →
, а вторая стабилизируется, x(t) →![]() , t→
, t→![]() . Наконец, если
. Наконец, если ![]() =
=![]() =
=![]() , то кроме неустойчивого узла (0,0) имеем линию стационарных точек – отрезок прямой
, то кроме неустойчивого узла (0,0) имеем линию стационарных точек – отрезок прямой 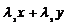 =
=![]() (рис. 9.3).
(рис. 9.3).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 |
