

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Если напротив, г субстрата меньше, чем биомассы, то необходимо прямое окисление субстрата молекулярным кислородом для понижения степени восстановленности углерода. В этих окислительных реакциях, катализируемых различными ферментами (оксидазами, оксигеназами и пероксидазами), не обнаружено никакого механизма улавливания энергии, т. е. в термодинамическом отношении они ведут к потере клеткой (рассеиванию в виде тепла) части свободной энергии субстрата, соответственно, энергетический выход падает. Таким образом, при существенном отличии г субстрата от г биомассы следует ожидать более низких энергетических выходов, чем при совпадении степени восстановленности углерода в субстрате и биомассе.
Химический состав биомассы. Элементный состав сухой биомассы бактерий по результатам статистической обработки многочисленных экспериментальных данных составляет: углерод − (46,2+2,4)%, водород − (6,8+0,4)%, кислород − (30,3+3,1)%, азот − (8,9+1,6)%, зола − 7,8%. Это соответствует энергосодержанию 4,2 ккал/г, кислородному эквиваленту 1,33кг О2/кг, степени восстановленности углерода 4,2 и среднему химическому составу: белки − 60%, нуклеиновые кислоты (ДНК и РНК) − 20%, углеводы − 15%, липиды − 5%. В определенных условиях может синтезироваться биомасса аномального состава. Например, при недостатке азота в среде происходит сверхсинтез полисахаридов, содержание которых в сухой биомассе может достигать 50 − 60%.
Затраты энергии на синтез различных компонентов биомассы неодинаковы: на синтез 1 кг липидов и полисахаридов требуется энергии в несколько раз меньше, чем на синтез белков и нуклеиновых кислот. Поэтому с увеличением в клетках весовой доли липидов, полисахаридов, а также поли−в−оксимасляной кислоты, энергетический выход возрастает.
Длина углеродной цепи молекул субстрата. Для транспортировки внутрь бактериальных клеток молекул с длинной углеродной цепью (высокомолекулярных) требуется их предварительное расщепление до мономеров с помощью внеклеточных ферментов (экзоферментов), т. е. возникают дополнительные энергетические затраты, энергетический выход биомассы снижается.
Соотношение затрат энергии на рост и поддержание жизнедеятельности. Процессы поддержания жизнедеятельности включают оборот клеточного материала (основной обмен), осмотическую работу по поддержанию градиентов концентрации веществ внутри и вне клетки, подвижность клеточных органелл и т. п. Энергетические затраты на поддержание жизнедеятельности увеличиваются в неоптимальных для роста условиях (рН, температура и т. д.), с увеличением сравнительной токсичности органических веществ и ростом концентрации компонентов, способных оказывать на микроорганизм токсичное действие. В соответствии со схемой 1.5 с ростом затрат субстрата на процессы поддержания жизнедеятельности энергетический выход падает.
Степень сопряженности конструктивного и энергетического обменов. Вся совокупность реакций биоокисления субстрата, идущих с выделением энергии (энергодающие процессы), называется энергетическим обменом веществ, а совокупность реакций синтеза, в которых энергия потребляется, − конструктивным обменом. Вопрос сопряженности конструктивного и энергетического обменов является одним из главных в энергетике роста микроорганизмов. Это сопряжение осуществляется через макроэргические соединения, основным из которых является АТФ. Энергия, выделяющаяся при окислении субстрата, может запасаться в АТФ, образующейся в результате фосфорилирования АДФ. Гидролиз АТФ приводит к выделению энергии, которая может обеспечивать выполнение различной работы: химической (биосинтез компонентов клетки), механической (движение клетки и её органелл), электрической (перенос заряда против градиента электрического потенциала), осмотической (перенос веществ против градиента их концентрации).
В аэробных условиях, когда перенос электронов от окисляемого вещества на кислород осуществляется по дыхательной цепи, энергетический обмен относительно независим от конструктивного обмена. В неоптимальных для роста микроорганизмов условиях происходит разобщение этих процессов путем окисления субстрата без образования АТФ и "холостого" гидролиза АТФ; энергия рассеивается в виде тепла, а не используется на физиологические нужды клетки. С увеличением степени сопряженности конструктивного и энергетического обменов энергетический выход биомассы возрастает.
Многочисленными исследованиями установлено, что при росте гетеротрофных бактерий на простых субстратах энергетический выход биомассы является физиологической константой (з = 0,6), если выполняются следующие условия: синтезируется биомасса нормального состава (содержание полисахаридов не более 20...30%), степень восстановленности углерода в субстрате и биомассе примерно одинаковы, степень сопряженности конструктивного и энергетического обменов максимальна, расход субстрата на поддержание жизнедеятельности значительно меньше его затрат на рост клеток.
Перечисленные условия, как правило, выполняются в системах неполной биологической очистки промстоков ЦБП, т. е. когда биоокислению подвергаются наиболее легкоокисляемые компоненты сточной воды, а самоокисление активного ила не получает заметного развития. При росте активного ила на трудноокисляемых компонентах промстоков энергетический выход биомассы ниже, в частности, на низкомолекулярных производных фенола максимальный энергетический выход составляет около 0,5, а на щелочном лигнине − не более 0,1 − 0,2.
Кислород в процессе аэробной биологической очистки в общем случае затрачивается на прямое химическое окисление и биоокисление. Для стоков ЦБП расход кислорода на прямое химическое окисление составляет не более нескольких процентов от общих его затрат.
Потребление кислорода на процесс нитрификации, ведущий к окислению аммонийного азота до нитритов и нитратов, также пренебрежимо мало. Поэтому уравнение энергетического баланса в кислородных единицах имеет вид:
Д ХПК сист. = Д О2; (1.10)
где Д ХПК сист. − изменение (уменьшение) ХПК системы "активный ил - сточная вода", кг О2 /м3, Д О2 − количество потребленного кислорода кг О2 /м3.
В процессе биологической очистки ХПК сточной воды снижается, а ХПК активного ила возрастает:
Д ХПК сист. = ДS − хx Д Х; (1.11)
где ДS − уменьшение ХПК сточной воды, кг О2 /м3, Д Х − прирост активного ила по сухому весу, кг/м3, хx − кислородный эквивалент (удельное ХПК) активного ила, кг О2 /кг.
Из (1.10) и (1.11) получим:
ДS = хx Д Х + Д О2; (1.12)
1 = з + Z; (1.13)
где Z = Д О2/ ДS − расход кислорода на 1 кг снятого ХПК сточной воды кг О2/ кг О2.
Соотношение (1.12), по существу, является уравнением материально-энергетического баланса: изменение ХПК сточной воды, пропорциональное изменению свободной энергии органических веществ, увязывается с приростом активного ила в единицах сухого веса.
Экономический коэффициент, равный приросту активного ила на 1 кг снятого ХПК промстоков, пропорционален энергетическому выходу:
у = ![]() ; (1.14)
; (1.14)
В системах неполной биологической очистки кислородный эквивалент активного ила достаточно стабилен. В среднем хxо = 1,33 кг О2 /кг (на единицу беззольной массы активного ила 1,42 кг О 2 /кг). Истинный экономический коэффициент уо, который равен экономическому коэффициенту, при условии, что скорость самоокисления биомассы и скорость потребления субстрата на поддержание жизнедеятельности пренебрежимо малы в сравнении со скоростью роста микроорганизмов, составляет
уо = ![]()
При исчерпании в среде легкоокисляемых компонентов часть микроорганизмов активного ила, лишившись доступного субстрата, переходит в эндогенную фазу (самоокисляется). Микроорганизмы, способные утилизировать трудноокисляемые компоненты, продолжают рост, но с более низким (чем при росте на легкоокисляемых веществах) энергетическим выходом биомассы. По этим причинам с повышением эффекта очистки энергетический выход падает и в пределе стремится к нулю. Экономический коэффициент также снижается, но в силу накопления в активном иле минеральных продуктов окисления (веществ с нулевым энергетическим уровнем) всегда больше нуля: у ≈ 0,1. Возможный диапазон изменения коэффициента Z, как следует из уравнения (1.13), составляет 0,4 − 1,0.
Прирост активного ила и потребление кислорода обычно относят к БПК5 сточной воды. Из (1.12) получим:
хx·Уl + Zl = ![]() ; (1.15)
; (1.15)
Уl = ![]() ; Zl =
; Zl = ![]() ; (1.16)
; (1.16)
Из уравнения (1.1) следует:
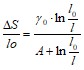 ; (1.17)
; (1.17)
В результате уравнение материально-энергетического баланса сводится к уравнению взаимосвязи прироста активного ила, потребления кислорода и глубины биологической очистки:
хx·Уl + Zl = 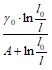 ; (1.18)
; (1.18)
1.4. КИНЕТИКА БИООКИСЛЕНИЯ В СИСТЕМАХ С АКТИВНЫМ ИЛОМ
Процесс биохимического окисления органических загрязнений в аэротенках является одним из промышленных микробиологических процессов, но имеет ряд особенностей.
Во-первых, активный ил − это не искусственный микробный ценоз, подобранный из чистых культур, а экологическая система микроорганизмов, самопроизвольно сформировавшаяся в результате ферментативной и экологической адаптации в реальных условиях процесса. Смена режима биологической очистки, в частности, изменение концентрации сточных вод, вызывает перестройку всего биоценоза. Длительность такого адаптационного перехода может составлять несколько суток и более. Естественно, что кинетика биоокисления будет различной в стационарном и переходном состояниях активного ила. Например, установлено, что для неадаптированного активного ила скорость биоокисления органических веществ промстоков прямопропорциональна концентрации ила. Наоборот, в адаптированных системах начальная скорость биоокисления прямопропорциональна концентрации сточной воды в широком диапазоне БПК, а для неадаптированных систем эта зависимость описывается кривой с насыщением.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 |
