
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
анатомическая схема путей кровотока в сердечно-сосудистой системе.
Артериальная часть системы содержит всего 15-20% общего объема крови и характеризуется высоким давлением. В центр схемы вынесена область транскапиллярного обмена, т. е. капиллярных (обменных) сосудов, на обеспечение оптимальной функции которых и направлена вся деятельность сердечнососудистой системы. Числе капилляров огромно (до 40 млрд.) в организме человека, площадь обменной поверхности капилляров очень велика - около 1000 м2, а площадь функционирующих капилляров составляет 200-350 м2 в покое. Вместе с тем, объем содержащейся в капиллярах крови в условиях покоя организма относительно невелики.
Как видно из схемы наибольшее количество крови содержится в области большого объема, которая содержит в 3- 4 раза больше крови, чем область высокого давления.
|
Сердечно-сосудистая система (функциональная схема) Цифры у названия органов – величина кровотока в покое (% к минутному объему); Цифры внизу – объем содержащейся крови (% к общему объему) в трех функционально различных областях сосудистого русла; вверху – название этих областей; стрелки – направление кровотока. |

СТРОЕНИЕ И КЛАССИФИКАЦИЯ СОСУДОВ МИКРОЦИРКУЛЯТОРНОГО РУСЛА
Долгое время основным критерием классификации мелких артерий, капилляров и вен служило изменение диаметра сосудов по мере их разветвления. Однако принадлежность микрососудов тому или иному звену микроциркуляторного русла устанавливается намного более точно, если учесть особенности строения стенки, распределение мышечных клеток, характеристики окружающих соединительнотканных компонентов, способы разветвления и соединения между собой микрососудов, участие их во внутриорганном распределении крови, а также в обменных процессах. Такого рода анализ привел к развитию фундаментальных представлений о строении связующего звена между артериями и венами, которое теперь называют микроциркуляторным руслом (Куприянов и др., 1975). Капилляры составляют главный компонент микроциркуляторного русла, но их функционирование стоит в сильной зависимости от кровотока в других микрососудах. Управление кровотоком и обменными процессами в органах и тканях осуществляется через посредство всей совокупности взаимосвязанных микрососудов.
Микроциркуляторное русло, несмотря на выраженные органные особенности, характеризуется определенной общностью плана строения (рис. 6.1). Как минимум, в этот план следует включить: а) комплекс микрососудов, состоящий из артериол, пре-капиллярных артериол, капилляров, посткапиллярных венул, венул и артериоло-венулярных анастомозов; б) последовательность их соединения в микрососудистых сетях; в) определенные количественные соотношения между микрососудами и их топографическую упорядоченность в тканях и органах. Наименования звеньев микроциркуляторного русла приведены в соответствии с Nomina histologica, принятой в 1970 г. на IX Международном конгрессе анатомов в Ленинграде. Наряду с данной морфологической классификацией микрососудов в литературе встречаются иные различные их наименования, что обусловлено большим разнообразием органных особенностей строения и функционирования микроциркуляторного русла. В табл. 6.1 представлены наиболее часто употребляемые в литературе термины и сделана попытка сопоставить их с морфологической классификацией микрососудов.
Капилляры. Эти сосуды являются центральным и главным звеном микроциркуляторного русла. Кровеносный капилляр представляет собой тонкостенную трубку; каждая боковая ветвь - это отдельный капилляр. По форме различают петлевидные капилляры с приводящим (артериальным) и отводящим (венозным) коленами; капиллярные клубочки; синусоидные капилляры, отличающиеся очень широким просветом; диффузную капиллярную сеть (подробнее см.: Куприянов и др., 1975).
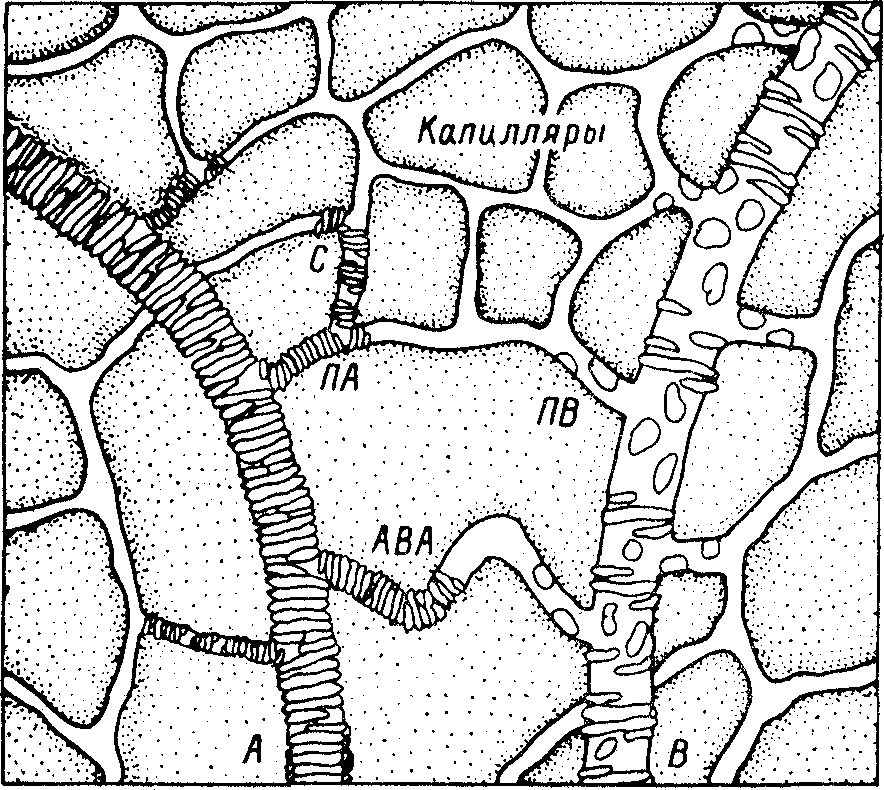
Рис. 6.1. Схема строения микропиркуляторного русла.
А - артериола, ПА - прекапиллярная артериола, ПВ - посткапиллярная венула, В - венула, С - прекапиллярный сфинктер, АВА - артериоло-венулярный анастомоз.
Диаметр капилляра колеблется от 4 до 20 мкм, в среднем составляя 7 - 8 мкм; в синусоидных капиллярах печени и костного мозга он может достигать 40-50 мкм.
Протяженность капилляров как в разных органах, так и в одном и том же органе является чрезвычайно вариабельной величиной - от 50 мкм до 1 мм (подробно сводку структурных параметров кровеносных капилляров см.: Шошенко, 1975). Здесь уместно отметить функциональное значение длины (и диаметра) капилляра, так как чем больше этот параметр, тем большее время кровь находится в нем. В органах капилляры образуют сети различной формы и сложности, конструкция которых приспособлена к местным особенностям кровотока. При построении сетей имеет место последовательное и параллельное соединение капилляров, в силу этого протяженность капиллярного звена от артериального до венозного конца может достигать значительной длины (например, в мышцах до 1.5 - 2 мм). За длину капилляра разные авторы принимают протяженность различных фрагментов капиллярной сети. Одни (например, Григорьева, 1954; Шошенко, 1975) за капилляр принимают сосуд от места его ответвления от артериолы до места его впадения в венулу; другие (Куприянов и др., 1975; Козлов и др., 1982) полагают, что в качестве капилляра следует рассматривать каждый фрагмент капиллярной сети между соседними узлами разветвления или слияния капилляров.
На венозном конце капилляры имеют больший диаметр (6 - 10 мкм), чем на артериальном (4 - 6 мкм); артериальный конец капилляра является самым узким сосудом.
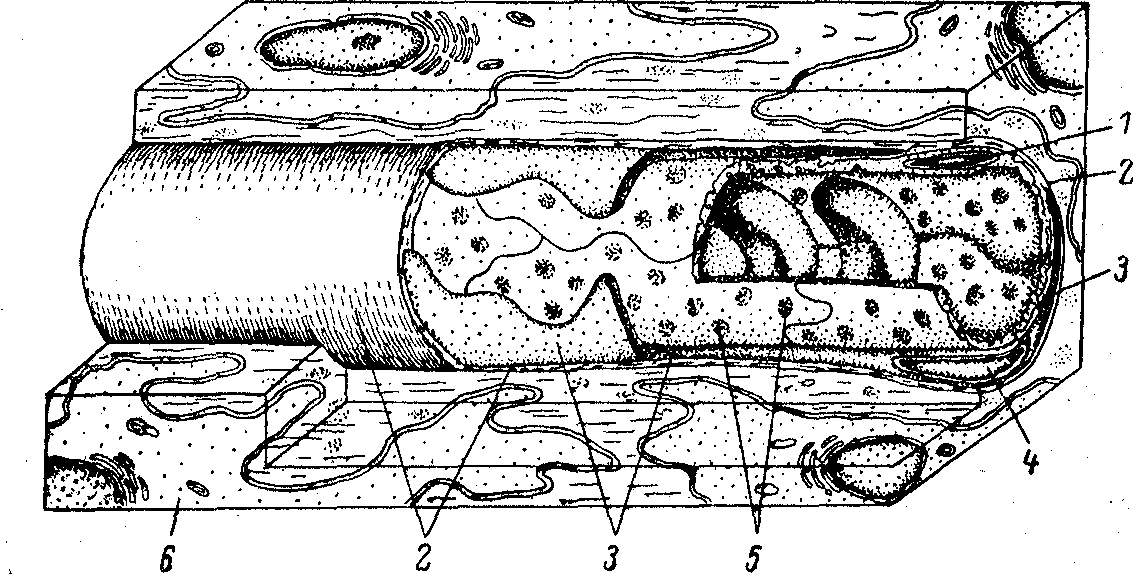
Рис. 6.2. Строение стенки кровеносного капилляра и его отношение к окружающей
ткани.
1 - эндотелиоцит, 2 - базальная мембрана, 3 - перицит, 4 - ядро перицита,
5 - микропиноцитозные везикулы, 6 - фибробласт.
Стенка капилляра образована эндотелиальными клетками, лежащими в один слой. Снаружи от них расположена базальная мембрана, в толще которой местами залегают отросчатые клетки - перициты (рис. 6.2). Роль последних достоверно не выяснена; высказано предположение об участии перицитов в регуляции роста эндотелиальных клеток. Показаны особенности топологических взаимосвязей перицитов и эндотелиальных клеток в различных органах, что делает вероятным представление о разной функции перицитов в них.
Действительно, перициты в стенке капилляров скелетной мышцы крысы, в отличие от таковых в миокарде, способны уменьшать свои размеры в ответ на воздействие вазоактивных веществ (Tilton et al., 1979). Отличительной особенностью эндотелиальных клеток капилляров является наличие многочисленных микропиноцитозных везикул, размеры которых варьируют от 50 до 90 нм. В истонченных участках эндотелиоцитов на долю везикул приходится до 40% объема клетки. Микропиноцитозные везикулы могут объединяться между собой в виде цепочек, образуя при этом каналы различной длины, которые в отдельных случаях могут насквозь пронизывать эндотелиальную клетку (Караганов и др., 1982). Везикулы, а также фенестры, которые представляют собой пронизывающие цитоплазму эндотелиальных клеток отверстия (диаметром 40- 60 нм), и стыки между эндотелиальными клетками участвуют в трансэндотелиальном переносе веществ.
Выявлен ультраструктурный фибриллярный субстрат эндотелиоцитов, обеспечивающий различные формы подвижности эндотелиоцитов (Hammersen, 1980). Несмотря на то что имеются прямые наблюдения, свидетельствующие о возможности набухания эндотелиоцитов, ведущего к перекрытию просвета капилляра (McCuskey, 1971), или об их сокращении (Weigelt et al., 1981), большинство авторов склонны рассматривать сократимость эндотелиоцитов в связи с проблемой проницаемости стенки капилляров, а не регуляции гемодинамики.
Как показали электронно-микроскопические наблюдения, следует различать три типа капилляров: соматический, висцеральный и синусоидный. Капилляры соматического типа характеризуются непрерывностью эндотелиального пласта и базальной мембраны. Стенка капилляров легко пропускает воду и растворенные в ней кристаллоиды, но малопроницаема для крупных молекул белка. Соматический тип капилляров обнаружен в коже, скелетных мышцах, миокарде, коре мозга. Для капилляров висцерального типа характерно наличие в эндотелии фенестр (окошек), вследствие чего подобный эндотелий нередко называют окончатым. Такой тип капилляров обнаруживается в тех органах, в которых секретируется и
всасывается большое количество воды с растворенными в ней веществами, как это наблюдается в железах, органах желудочно-кишечного тракта, почках. Фенестрированный эндотелий характерен также для эндокринных органов, в которых происходит быстрый трансэндотелиальный транспорт макромолекул. В капиллярах синусоидного типа, обнаруженных в костном мозге, селезенке, печени, эндотелиальные клетки отделены друг от друга различимыми щелями, в области которых обычно отсутствует и базальная мембрана. Такой прерывистый тип эндотелия обусловливает высокую проницаемость стенки капилляров не только по отношению к белковым молекулам, но и к клеткам крови. Подробное описание субмикроскопического строения капилляров можно найти в специальных работах (Шахламов, 1971; Куприянов и др., 1975; Чернух и др., 1975; Караганов и др., 1982).
Несмотря на небольшую толщину стенки капилляров, их способность к растяжению мала. Это определяется не столько величиной модуля Юнга материалов стенки, который для базалъной мембраны капилляров равен 102 Н м-2, что близко к аналогичной величине для коллагена, сколько механическими свойствами окружающей капилляры соединительной ткани.
Прямые измерения показывают, что окружающая капилляры ткань обусловливает 99% их суммарной жесткости. С точки зрения механики предложено даже рассматривать капилляр не как трубку, а как туннель в геле (Fang, Zweifach, 1971). Растяжимость такого капилляра - туннеля, как показывают расчеты, составляет примерно 1/3 от растяжимости капилляра-трубки.
Прекапиллярный сфинктер. Некоторые капилляры в местах своего ответвления от артериол имеют прекапиллярный сфинктер. Он состоит обычно из 1 - 2 гладких мышечных клеток, кольцом охватывающих устьевой отдел капилляра. При сокращении гладких миоцитов происходит сужение или перекрытие просвета капилляра, тем самым регулируется
поступление крови в капиллярное русло. Однако прекапиллярные сфинктеры обнаружены не во всех органах; так, не доказано их существование в скелетных мышцах. Высказываются сомнения относительно участия именно прекапиллярных сфинктеров в регуляции капиллярного кровотока, хотя несомненно прекапиллярная артериола выполняет роль регулятора, контролирующего кровенаполнение капилляров. Поэтому в физиологической литературе понятие прекапиллярного сфинктера порой используется не в строгом морфологическом смысле, а в функциональном (разд. 6.2).
Описаны явления спонтанной временной закупорки устьевых отделов капилляров крупными ригидными клетками белой крови, симулирующие проявление "вазомоции" прекапиллярных сфинктеров. Хотя прекапиллярные сфинктеры и контролируют кровоток в соответствующих участках капиллярных сетей, однако, поскольку активация отдельных сфинктеров не синхронизирована, их не включают в число регуляторов общего периферического сопротивления органного русла, влияющих на уровень системного кровяного давления (Wiedeman et al., 1976).
Артериолы. Образуя пути доставки крови в капиллярное русло, артериолы тем самым являются связующим звеном между артериями и капиллярами. В зависимости от особенностей строения и функционирования различают собственно артериолы, ближе расположенные к артериям (их диаметр в среднем составляет 20 - 35 мкм), и прекапиллярные артериолы (диаметром 12 - 15 мкм), по которым кровь поступает непосредственно в капилляры.
Характерной чертой строения артериол является наличие в составе их стенки гладких мышечных клеток, расположенных, в отличие от артерий, только в один ряд. По мере приближения к капиллярам и перехода артериол в прекапиллярные артериолы, слой гладких миоцитов перестает быть сплошным, т. е. их относительное количество в стенке сосуда уменьшается.
Обычно артериолы сопровождаются венулами одноименных порядков. В отличие от них прекапиллярные артериолы, как правило, следуют на некотором расстоянии от одноименных посткапиллярных венул и отделены от них сетью капилляров. Артериолы, так же как и внутриорганные артерии, обозначаются по порядкам ветвления, нумерация которых ведется от проксимальных узлов ветвления к дистальным.
В артериолах эндотелиальные клетки плотно прилежат друг к другу. Снаружи от них лежит базальная мембрана. Гладкие миоциты располагаются либо циркулярно, либо по геликоиду, при этом одна гладкомышечная клетка может до 1.5 раза обернуться вокруг сосуда. При циркулярном расположении мышечных клеток угол их наклона к продольной оси составляет около 90°. По мере приближения к капиллярам слой гладких миоцитов становится более разреженным; угол их наклона к продольной оси сосуда порой уменьшается, при этом в стенке сосуда формируется как бы своеобразная пружина, которая при сокращении миоцитов должна сжиматься. Расчеты показывают, что если угол наклона гладких миоцитов к продольной оси сосуда много меньше 45°, то при их сокращении артериола должна делаться короче и шире. При сокращении обычно расположенных (циркулярно) гладких миоцитов просвет сосуда суживается.
В стенке артериол обнаружены специальные структуры - миоэндотелиальные контакты, образованные выростами (протрузиями) эндотелиальных клеток, которые проникают через базальную мембрану и вступают в контакт с миоцитами. Функциональное значение миоэндотелиальных контактов не раскрыто. Возможно, они являются специализированными образованиями, передающими действие приносимых с кровью вазоактивных веществ на миоциты. По мере приближения к капиллярам число миоэндотелиальных контактов в стенке сосудов возрастает; наибольшее их количество наблюдается в области прекапиллярных сфинктеров (Rhodin, 1967). Снаружи от гладких миоцитов расположены в небольшом числе фиброциты, эластические и коллагеновые волокна. Примерные соотношения между разными структурными компонентами в стенке артериол приведены в табл. 6.1.
К числу органоспецифических особенностей относятся способ и выраженность ветвления артериол. Обычно артериолы по своему ходу отдают одиночные боковые ветви; наряду с этим встречаются дихотомический и реже трихотомический типы ветвления артериол. Число порядков ветвления артериол, или количество генераций сосудов, образуемых ими до перехода в капилляры, значительно варьирует в разных органах. В крыле летучей мыши от главной питающей артерии, диаметр которой равен 97 мкм, до капилляров имеется 4 генерации микрососудов (Mayrovitz et al., 1975); в стенке тонкой кишки крысы - 5 генераций (Bohlen, Core, 1977). В мышце, подвешивающей яичко (m. cremaster), у крысы между вторичными ветвями питающей артерии и метартериолами насчитывается 4 генерации артериол (Baez, 1973); в мышце напрягателя фасции бедра (m. tenuissimus) у кошки артериолы, отходящие от центральной питающей артерии, средний диаметр которой составляет 72 мкм, дают ветвления 11 порядков до капилляров (Myrhage, Eriksson, 1980). В брыжейке кошки артериолы диаметром 60 мкм, отходящие от кишечных артерий, образуют 4 порядка ветвления (Gaehtgens, 1971). Число порядков ветвления артериол, равно как и тип ветвления, имеют самое непосредственное отношение к распределению давления в микрососудистых сетях.
Принято различать два типа сетей микрососудов. Для одного из них характерно последовательное дихотомическое разделение артериол вплоть до образования капиллярной сети; при этом анастомозирование между соседними микрососудами осуществляется на уровне капилляров. При таком последовательном типе ветвления артериол (Zweifach, Lipowsky, 1978) происходит постепенное падение давления по руслу. Подобный способ ветвления артериол обнаружен в коже, сальнике, скелетных мышцах. Другой тип, получивший название аркадного, обусловлен анастомозированием артериол. Аркадный тип строения микроциркуляторного русла и особенности кровотока в нем достаточно подробно описаны в брыжейке тонкой кишки, в крыле летучей мыши (Nicoll, 1969) и в ряде других органов. При аркадном построении сети артериол наблюдается ступенчатое падение давления на путях притока крови в капиллярное русло, т. е. вся генерация сосудов, отходящих от аркадной (анастомозирующей) артериолы, имеет одно и то
же входное давление.
Венулы. Пути оттока из микроциркуляторного русла образуются посредством слияния нескольких капилляров, в результате чего формируются посткапиллярные венулы. Их диаметр очень вариабелен, в среднем составляет 15-20 мкм. По ходу посткапиллярных венул в них впадают другие капилляры, эти венулы нередко называют собирательными. Из посткапиллярных венул, для которых различают несколько порядков слияния, кровь поступает в более крупные венулы - диаметром 30-50 мкм.
Характерной особенностью строения стенки посткапиллярных венул является большое число перицитов, расположенных снаружи от базальной мембраны почти сплошным слоем. Стенка посткапиллярных венул легко растяжима и обладает сравнительно высокой проницаемостью. Эндотелиальные клетки представляются уплощенными, среди них встречаются фенестрированные. Гладкие мышечные клетки в посткапиллярных венулах отсутствуют; они появляются в стенке собственно венул (табл. 6.1). В венулах диаметром 50-100 мкм эндотелий и базальная мембрана окружены одним сплошным слоем гладких миоцитов. Снаружи от них лежат фибробласты и коллагеновые волокна, образующие адвентициаль-ную оболочку, толщина которой по мере увеличения диаметра венул нарастает.
Порядки слияния венулярных сосудов в целом повторяют порядки ветвления артериол. Однако в области посткапиллярных венул значительно выражено анастомозирование между их притоками, что придает начальным звеньям венулярного русла вид мелкоячеистой сети. В венулах имеются клапаны, расположение которых определяет направленность движения крови на путях оттока.
Артериоло-венулярные (или артерио-венозные) анастомозы. Эти структуры давно известны как короткие сообщения между артериолами и венулами, через которые часть крови непосредственно из артериального звена поступает в венозное, минуя основную сеть капилляров (рис. 6.1, 6.3). Доказано участие кожных артериоло-венулярных анастомозов в терморегуляции. Вместе с тем повсеместное распространение артериоло-венулярных анастомозов и обусловленное их участием образование путей шунтирующего кровотока имеет весьма важное функциональное значение.
Диаметр артериоло-венулярных анастомозов обычно колеблется от 15 до 100 мкм, хотя встречаются анастомозы и с большим просветом. Вообще диаметр и длина анастомоза зависят от уровня тех сосудов, между которыми он замыкает связь. В артериальном сегменте артериоло-венулярного анастомоза имеется скопление гладких миоцитов, регулирующих степень его раскрытия. Детальное изучение морфологии артериоло-венулярных анастомозов позволило выделить среди них шунты и полушунты. В структурах типа шунтов наблюдается резкий переход артериального сегмента в венозный. Полушунты отличаются тем, что на своем протяжении имеют промежуточный сегмент, который по характеру строения стенки сходен с капилляром.
ЛИТЕРАТУРА
Физиология кровообращения. Физиология сосудистой системы. Под. Ткаченко. Ленинград, Наука, 1984 г., 652 с.
Капиллярное кровообращение. Тбилиси: Изд. АН ГрузССР,
1958г. 110 с.
, , Микроциркуляция. Москва, Медицина, 1975 г.,153 с.
Кровеносные капилляры. Новосибирск: Наука, 1975. 372 с.
