
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
![]()
ии нормальных скелетных мышц.
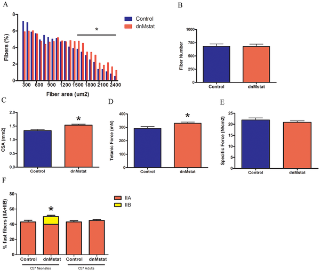
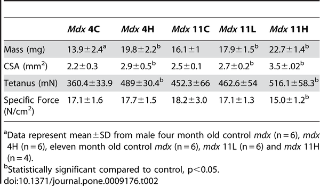
Чтобы выяснить влияние торможения миостатина в камбаловидной мышце, было оценено распределение волокон в C57 Bl / 6 группах. Не существовало никакого влияния dnMstat в камбаловидной мышце на тип волокон в C57 группе взрослых (рис. 3F). Однако произошел сдвиг в сторону быстрых типов волокон в C57 группе новорожденных, до 10% обработанных волокон мышц были MHC IIB типа, в то время как нет волокон типа IIB обнаруженных в контрольных группах. MDX группы демонстрируют гиперэкспрессию dnMstat, повышенную массу тела по сравнению с контрольной группой (рис. 4, C, D).Увеличение размера скелетных мышц конечности было сопоставимо с результатами ранее описанных подходов к торможению миостатина. В MDX 4H группе масса передней большеберцовой, икроножной и четырехглавой мышцы бедра у обработанных животных была на 38%, 34% и 46% больше, чем в контрольной, соответственно (рис. 4в).Увеличение мышечной массы, наблюдаемое в этих мышцах, сохраняется до одиннадцати месяцев в MDX группе 11H. В то время как различия веса 11L и 11H MDX мышц не были статистически различны, но были больше, чем вес контроля и была заметная тенденция к большей величине роста мышц, когда торможение миостатина было начато в возрасте одного месяца (MDX 11H) по сравнению с неонатальным периодом (MDX 11L) (рис. 4).Вес сердца мышей MDX увеличился на 25% в группе MDX 4H и 22-42% в MDX 11L и 11H группах (рис. 4, D).Отношение веса сердце / тело (мг / г) не изменится в MDX 4H группах (4 ± 0,1 против 4 ± 0,3). Группа MDX 11L имела соотношение 4,2 ± 0,1, что меньше, чем в контрольной. Это, скорее всего, потому, что увеличение массы тела в этой группе немного опережало увеличение
![]()
веса сердца.
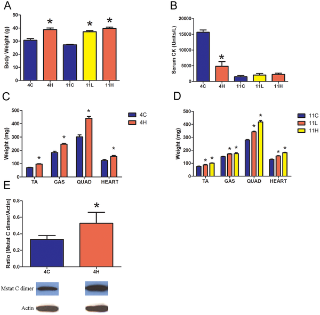
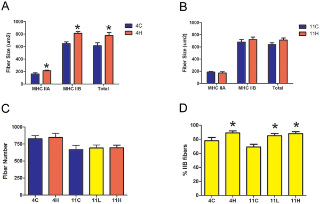
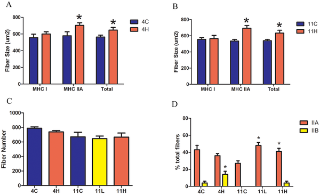
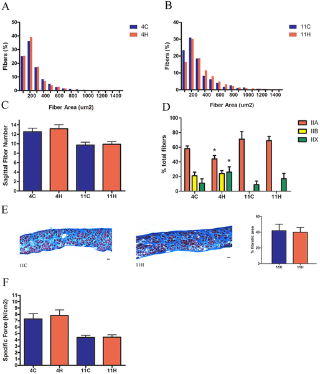
Сывороточная креатинкиназа (CK) является мерой состояния скелетных мышц в организме в целом. У MDX мышей и людей страдающих от мышечной дистрофии Дюшенна отсутствие дистрофина приводит к повреждению сарколеммы и повышению проницаемости мембран, и тем самым подъем сывороточной КФК. Торможение миостатина привело к значительному снижению в сыворотке крови уровня КФК в группе MDX 4H по сравнению с контрольной MDX группой (рис. 4В), однако сывороточная КФК у MDX 11L и 11H групп не отличалась от одиннадцати месячных MDX мышей, получавших лечение. Иммуногистохимия была использована для измерения площади поперечного сечения (CSA) и оценки тяжелой цепи миозина (МНС)в составе мышечных волокон из EDL, камбаловидной мышцы и диафрагмы MDX групп. Эти скелетные мышцы были выбраны для анализа, чтобы определить, есть ли группы мышц с различными типами волокон и прогрессирование заболевания дифференциально реагирует на торможение миостатина. EDL представляет собой прототип "быстрых" мышц, состоящих в основном из быстрых гликолитических MHC IIB волокон, а камбаловидная мышца представляет собой прототип "медленных" мышц и заполняется медленными окислительными MHC I волоконами. Диафрагма содержит смешанный тип распределения волокон. В мышцах конечностей наблюдалось увеличение мышечной массы волокон без изменения числа волокон, за исключением EDL в группе MDX 11H, которые не подверглись гипертрофии (рис. 5, 6). Размеры волокна были дополнительно классифицированы в соответствии с типом MHC чтобы определить, какой тип волокна приходится на общую гипертрофию. В соответствии с результатами предшествующих исследований, которые обнаружили, что миостатин преимущественно действует на быстрый тип волокон, средний CSA тип IIA и IIB волокон увеличился в EDL в то время как средняя CSA только из волокон типа IIA была увеличена в камбаловидной мышце(рис. 5А, B и 6А, Б). EDL была на 11% больше волокон типа IIB в группе MDX 4H и на 19% больше типа IIB волокон в MDX 11H группе (рис. 5). Кроме того, в камбаловидной мышце было на 13% больше волокон типа IIB в группе MDX 4H и на 20% больше типа IIA волокон в MDX 11H группе (рис. 6). В обоих мышцах конечностей не было существенных различий между MDX 11L и 11H группами. На обоих концах не было никаких различий в размерах, распределении волокон или количестве сагиттальной клетчатки в диафрагме (рис. 7A, B, C). В диафрагме наблюдалось увеличение IIx типа волокон и снижение волокон типа IIA у MDX 4H групп (рис. 7). Диафрагма MDX 11H была также проанализирована трехцветным окрашиванием Masson, чтобы оценить степень инфильтрации соединительной ткани. Как показано на рис. 7E, доля фиброза в диафрагме была неизменной на длительный срок сверхэкспрессии dnM
![]()
stat.
![]()
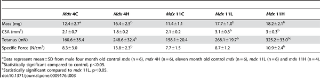
Функции EDL, камбаловидной мышцы и диафрагмы были оценены, чтобы определить, избыточную экспрессию dnMstat, повышенную прочность мышц. MDX 4H в EDL увеличилось абсолютное производство тетанической силы, сравнимое с увеличением CSA в результате изменений силы (табл. 2). В отличие от MDX 4H в камбаловидной мышцы генерируется 55% больше тетанической силы и на 66% более высокую удельную силу, а сила в диафрагме не изменилась (табл. 3, рис. 7E). Эти данные указывают на короткий срок блокады миостатина. В одиннадцать месяцев камбаловидная мышца продолжала демонстрировать повышенную абсолютную и удельную силу, и эффект был более выражен в MDX 11H группе (табл. 3). В то время как CSA был увеличен в MDX 11L, тетаническое сила не изменились (табл. 2). Абсолютная сила EDL был слегка увеличена в группе MDX 11H, но удельная сила снизилась на 21%. Диафрагма MDX 11H не продемонстрировала никакой разницы в силе (рис. 7E). Таким образом, долгосрочное ингибирование миостатина привело к значительному улучшению производительности силы только в камбаловидной мышце.
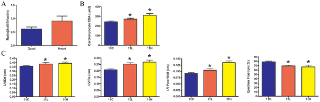
Для изучения взаимодействия между миостатином и инсулиноподобным фактором роста-I (IGF-I) гомогенизаты четырехглавой мышцы из MDX 4H группы были проанализированы с помощью иммуноблоттинга на наличие миостатина, и ИФА - на содержание IGF-I. Существовало примерно на 60% больше миостатина C-димера в обработанных мышцах (рис. 4E), несмотря на отсутствие разницы в IGF-I содержании. Функциональное действие торможение миостатина на дистрофическое сердце было определено с помощью эхокардиографии. Лечение MDX сердца и четырехглавой мышцы в течение одинадцати месяцев были исследованы на уровне рецепторов активина IIB. Белок и активин IIB рецепторы в этих тканях были одинаковыми (рис. 8а).Дозы ингибиторов миостатина зависимых от увеличения сырой массы сердца (рис. 4C, 4D, 8Б). В группе MDX 11H фракция выброса была сокращена на 11%, а толщина стенки левого желудочка составляет на 43% толще (рис. 8в).
![]()
Стойкое ингибирование миостатина через AAV опосредованную экспрессию нового ингибитора миостатина в печени приводит к увеличению массы скелетных мышц в норме и у MDX мышей. Важно отметить, что не только секреция в печени в результате широкого ингибирования миостатина, но она также дает стабильность в течение длительного периода времени, который не может быть возможен при доставке в скелетные мышцы. Доза ингибитора сокращенно "L" для низких доз и "H" для высокой дозы. В зависимости от дозы, масса скелетных мышц была увеличена в MDX 11L и 11H группах. Прибыль скелетной мышечной массы в нашем исследовании сопоставимы с введением нейтрализующих антител против миостатина или пропептида, и превосходит результаты предварительного исследования AAV опосредовано системного ингибирования миостатина. Поскольку данные свидетельствуют о том, что миостатин действует на клетки-сателлиты, так и взрослые волокна, увеличение мышечной массы может произойти через гипертрофию существующих мышечных волокон, или через образование новых волокон (гиперплазия). В предыдущих исследованиях, удаление миостатина генной терапией способствует как гипертрофии и гиперплазии скелетных мышц, в то время как послеродовое ингибирование миостатина индуцирует только гипертрофию. Двойной механизм повышения мышечной массы был также очевиден у MDX мышей на протяжении всей жизни. В текущем исследовании, во всех мышцах мышечный рост был обусловлен гипертрофией, а не гиперплазией. Этот вывод подтверждает мнение, что пренатальное отсутствие миостатина приводит как к гипертрофии, так и гиперплазии в скелетных мышцах, в то время как послеродовое торможение миостатина способствует росту мышц, прежде всего гипертрофии.
Однако, когда анти-миостатиновые антитела вводились два месяца мышам тяжелым комбинированным иммунодефицитом или миостатин был постнатально инактивирован использованием Cre-LoxP системы, у четырех месячных мышей не было никаких изменений в MHC профиле камбаловидной и EDL мышц. Таким образом, это первый доклад о торможении миостатина, которое приводит к изменению MHC содержания в послеродовых скелетных мышцах. Причины расхождений между нашими выводами у C57 Bl / 6 животных и предыдущих докладов могут быть связаны с эффективностью метода ингибирования или возраста животных, когда торможение миостатина было начато. Примечательно, что произошел сдвиг в сторону быстрых MHC изоформ в камбаловидной мышце в C57 группе новорожденных, но не в C57 группе взрослых. Эти результаты предполагают, что существует критический период развития мышц до трехмесячного возраста в течение которого миостатин можно регулировать после рождения. Типирование волокон у мышей MDX 1 месяца в камбаловидной мышце показали 30% волокон типа 1 и 70% волокна типа IIA, что согласуется с предыдущими докладами. Отсутствие типа IIB волокон на начало торможения миостатина в возрасте одного месяца предлагает новую роль миостатина в спецификации типа волокна. Преимущество селективного выживания быстрых типов волокон, как представляется, может активно переключаться на быстрые MHC изоформы, которые, вероятно, опосредовано регулируются механизмом, действующим на существующие волокна, потому что число волокон не изменилось. Потенциальным посредником в данном процессе является наличие большой активной популяции спутников. Быстрый постнатальный роста мышц у C57 Bl / 6 животных частично обусловлен слиянием клеток-сателлитов существующих мышечных волокон, а у MDX животных есть в большом масштабе фазы дегенерации - регенерации в возрасте 4-6 недель. Наблюдение типа переключения волокон у новорожденных мышей C57 и у всех MDX животных при активации однородного бассейна спутниковых клеток может внести вклад в процесс спецификации волокон под влиянием миостатина. Другой возможностью является то, что могут быть отдельные "медленные" и "быстрые" субпопуляции клеток, и спутниковая селективная активация «быстрой» субпопуляции при торможении миостатина может быть заполнение скелетных мышц быстрым типов волокон. Кроме того, наблюдаемые изменения в экспрессии MHC изоформ может быть связана с прямым действием на мышечные волокна, а не включать спутниковые клетки. Дальнейшие эксперименты необходимы, чтобы очертить механизмы и точные сроки типа переключения волокон нормальных скелетных мышц, индуцированных торможением миостатина.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
