
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Системное ингибирование миостатина передачей генов у нормальных и дистрофических мышей
Введение
Миостатин или фактор 8 (GDF-8) дифференциации является членом семьи трансформирующих ростовых факторов ? и является мощным негативным регулятором мышечной массы. Миостатин высоко сохраняется у разных видов и функционально инактивированный приводит к увеличениию мышечной массы у коров, собак, овец, мышей и человека. Наоборот, внематочная избыточная экспрессия миостатина вызывает атрофию скелетных мышц. Множество экспериментов показали, что отсутствие миостатина из-за генетических нарушений или торможение приводит к увеличению мышечной массы. В то время как эффект ингибирования миостатина на скелетные мышцы зависит от сроков и механизма блокады, всегда есть увеличение мышечной массы за счет роста мышечных волокон (гипертрофии) и / или увеличение количества волокон (гиперплазия). Постнатальное ингибирование миостатина достигается путем нейтрализирующих антител, направленных против миостатина, вводимых нормальных C57 Bl / 6 мышей и MDX мышам, что приводит к росту мышц за счет гипертрофии и гиперплазии. Общей чертой мышиных моделей является компенсаторная регенерация мышечных волокон за счет активации спутниковых клеток, пролиферации и слияния с существующими мышечными волокнами или формирования мышечных волокон De Novo. Поэтому данная терапия весьма перспективна для лечения мышечной дистрофии из-за ее способности увеличивать размер волокна, повысить регенерацию мышц и регулировать фиброз. Торможение миостатина привело к значительному улучшению дистрофической патологии у модели MDX, смешанные результаты у тяжелых дистрофических моделей (DSG) или никакого улучшения у тяжелых дистрофических моделей (GSG, ду). Это торможение миостатина не улучшает тяжелой мышечной дистрофии, что не удивительно, поскольку миостатин не приводит к устранению генетического дефекта. Для мышечной дистрофии торможение миостатина может быть использовано для увеличения коррекции первичного дефекта путем краткосрочной прибыли мышечной массы. Увеличение мышечной массы может защитить от повреждения, вызванного сокращением при уменьшении количества мышечных волокон, чтобы произвести определенное количество силы. Модуляция миостатина также имеет терапевтический потенциал для болезненных состояний, которые связаны с потерей нормальной мышечной массы, таких как кахексия, атрофия, саркопения. Тем не менее, существует возможность того, что торможение миостатина в сердце может помешать сердечной адаптации к основной сердечной болезни, которые могут возникнуть при старении, и мышечной дистрофии. Предыдущие доклинические исследования у MDX мышей - модели мышечной дистрофии Дюшенна, ориентированные на миостатин, использовали нейтрализующие антитела, инъекции миостатина пропептида и рекомбинантный адено-связанный вирус (ААВ) для опосредованной экспрессии ингибитора миостатина из нескольких тканей. Эти исследования показали, что все скелетные мышцы имели улучшение, хотя животные были исследованы в течение 3-4 месяцев после начала лечения. При мышечной дистрофии в результате прогрессивного патологии скелетных мышц, долгосрочные исследования имеют решающее значение для определения того, как предлагаемое лечение будет исправить патологию с течением времени и оценить эффект ингибирования миостатина на функцию сердца. В возрасте девяти месяцев MDX мыши сначала имеют признаки прогрессивной дилатационной кардиомиопатии, которые не были проанализированы в этих исследованиях. Цяо и соавт. недавно сообщалось, что экспрессия AAV опосредованного пропептида миостатина увеличивает размер мышц и улучшает патологию у животных MDX. В то время как они предположили, что это было связано с секрецией печенью, иммуноблоттинг не показывают экспрессии пропептида в печени и ОТ-ПЦР показал экспрессию трансгена в печени. Не исключено, что пропептид не экспрессируется в печени, но и в сердце, а также других неэкранированных скелетных мышцах. Haidet и соавт. использовали внутримышечные инъекции AAV для сверхэкспрессии эндогенных белков. Ограничением данного исследования является то, что ни один из ингибиторов не является специфичным для миостатина. Например, фолистатин как родственный ген (FLRG) связываются с активином в дополнение к миостатину и GDF связанному белку сыворотки 1 (GASP1). Наблюдаемое благотворное влияние на рост мышц может зависеть от сигнальных путей, кроме миостатина, которые не являются специфическим для мышц, и модуляция без особенностей физиологии мышечной ткани может ограничить клиническое применение. В то время появился значительный интерес к использованию торможения миостатина для улучшения мышечной дистрофии и других заболеваний, хотя всеобъемлющие долгосрочные исследования постнатального ингибирования миостатина еще не проводились. Оценки, включая оценку функционирования медленных и быстрых типов мышц, а также диафрагмы и сердца не достаточно. Диафрагма наиболее сильно имеет дистрофические изменения у мышей MDX. Прямое введение во все эти ткани не представляется возможным и дистрофические мышцы не поддерживает долгосрочные трансдукции AAV без стабилизации мембран. Потеря трансдукции AAV с течением времени, скорее всего, за счет оборота мышечных волокон, о чем свидетельствует потеря AAV опосредованной экспрессии IGF после четырех месяцев в мышцах MDX (неопубликованные наблюдения ER Бартон и HL Sweeney) и аналогичные потери AAV опосредованного пропептида миостатина. Для решения этих вопросов мы разработали метод, который позволяет достичь стойкого ингибирования миостатина одной дозой AAV при сверхэкспрессии новых ингибиторов миостатина из печени у C57 Bl / 6 и MDX мышей. C57 Bl / 6 мыши рассматриваться как новорожденные или молодые люди и исследуются через три месяца после инъекции, чтобы определить эффект ингибирования миостатина на мышечную массу у нормальных животных. Mdx мышам, новорожденным или в возрасте одного месяца, вводили препарат, их исследовали в четыре и одиннадцать месяцев, чтобы оценить эффект торможения миостатина на ранних и поздних стадиях мышечной патологии. Функциональная и гистологическая оценка камбаловидной мышцы, длинного разгибателя пальцев (EDL), диафрагмы и сердца проводилась чтобы определить системный эффект ингибирования миостатина. Эти выводы имеют важное значение для будущей терапии, которые используют пути и регулирующую роль миостатина в скелетных и сердечной мышцах.
Результаты
Чтобы экспрессировать стабильные циркулирующие ингибиторо миостатина, доминантный негативный пропептид миостатина был спроектирован (dnMstat), в паре с промотором ( LSP) и AAV псевдотипом 2/8 LSP. dnMstat. AAV псевдотип 2/8 имеет тропизм к печени и после внутривенной инъекции демонстрирует превосходную трансдукции в печени по сравнению с другими серотипами AAV. Экспериментальный дизайн исследования показан на рисунке 1а. C57 Bl / 6 мыши взяты в периоде новорожденности (п = 6 контроль, п = 6 основная C57 новорожденные), либо в три месяца (п = 6 контроль, п = 6 основная C57 взрослые) и проанализированы через три месяца после вирусной инъекции. MDX мыши были использованы для изучения влияния торможения миостатина на прогрессирование мышечной дистрофии. Уровни dnMstat были оценены в сыворотке крови всех групп MDX (рис. 1E). В возрасте четырех месяцев было ~ 3 кратное увеличение циркулирующих ингибиторов. Разница в циркуляции ингибитора поддерживается на протяжении всего исследования. Вирусная инъекция неонатальным животным эффективно привела к снижению дозы dnMstat, в то время как вирусные инъекции у мышей одного месяца требуют высоких доз dnMstat. В целях упрощения обсуждения, мы назвали экспериментальные группы с учетом возраста животного и дозы ингибитора. Небольшой группе новорожденных мышей MDX вводили препарат, а затем их исследовали в возрасте четырех месяцев, чтобы определить влияние возраста на вирусную экспрессию (п = 4 лечиться, MDX обозначается 4L). За одиннадцать месяцев мыши рассматриваться как новорожденные (п = 6, обозначается MDX 11L) или в возрасте одного месяца (п = 4, обозначается MDX 11H) с общим набором элементов лечения (п = 6, обознача
![]()
ется MDX 11C).
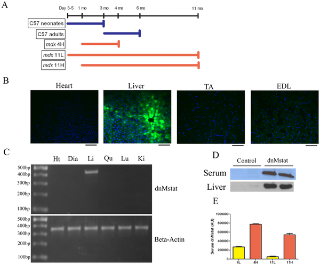
Специфика LSP была подтверждена внутривенной доставкой 1E12 копий генома AAV2 / 8 LSP. GFP. Экспрессия GFP была обнаружена исключительно в печени, и флуоресценция не наблюдается в сердце или скелетных мышцах (рис. 1б).Распределение в тканях экспрессии трансгена в дальнейшем оценивается ОТ-ПЦР в шести различных тканях. Экспрессия трансгенов была обнаружена только в печени (рис. 1в). AAV2 / 8 LSP. dnMstat вводили взрослым C57 Bl / 6 мышам и высокий уровень ингибиторов был обнаружен в крови, а также в печени после одной недели (рис. 1D). C57 Bl / 6 животных умерщвляли через неделю после вирусной инъекции для оценки острых изменениий в деятельности ключевых внутриклеточных регуляторов роста мышц. В рассматримой четырехглавой мышце произошло снижение фосфорилированной Smad 2/3, снижение фосфорилированных JNK и увеличение фосфорилированных Akt (п = 4) (рис. 2). Эти результаты показывают, что печень была жизнеспособной мишенью для AAV опосредованной сверхэкспрессии циркулирующих ингибиторов миостатина.

Эффективность ингибирования миостатина dnMstat была оценена путем анализа размера мышц и их функции. Экспрессия dnMstat привела к 14-22% увеличению мышечной массы четырехглавой мышцы, передней большеберцовой мышцы и длинных разгибателей пальцев (EDL) в C57 группе взрослых, в то время как 32-43% прироста в этих мышцах наблюдалось в группе C57 новорожденных (табл. 1). Таким образом, более выраженное увеличение мышечной массы наблюдалось у не-дистрофических животных, когда торможение было начато в начале жизни. Вес камбаловидной мышци и сердца не изменился в обеих C57 Bl / 6 группах. В C57 группе взрослых EDL прошли гипертрофию без гиперплазии, как показано сдвиг вправо без изменения числа волокон (рис. 3а, б).EDL стала ?? примерно на 15% больше по площади поперечного сечения (CSA) и силы (рис. 3, D). Взятые вместе эти данные свидетельствуют о экспрессии dnMstat, которая привела к увеличению мышечной массы, силы и гипертрофии без гиперплаз

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
