
Количественный метод полимеразной цепной реакции
ДНК была извлечена из мышц реципиентов путем инкубации в 400 мкл TE / SDS буфера (100 мМ Трис, рН 8, 1 мМ ЭДТА, 0,5% SDS) с добавилением протеиназы К (400 мкг, Roche Products Pty Ltd, Dee Why, Австралия, http://www. ) при 56 ° С в течение ночи при перемешивании. После обработки, 75 мкл NH4OAc и 1 мл абсолютного этанола перемешивали, затем центрифугировали в течение 20 минут. Супернатант удаляли, а осадок ДНК промывают в 70% этаноле (1 мл) и центрифугируют в течение 10 минут. ДНК сушат на воздухе и вновь растворяют в Milli-Q воды (100 мкл) и хранят при 4 ° C. Количественная обратная полимеразная цепная реакция (ПЦР) проводится с использованием праймеров в следующей последовательности: Y хромосомы вперед (TGG AGA GCC ACA AGC ТАА CCA), Y хромосоме обратно (TCC CAG CAT GAG AAA GAT TCT TC). Глицеральдегид-3-фосфатдегидрогеназа (GAPDH) была использована для количественной оценки общего количества ДНК, присутствующих в образцах со следующей последовательностью: GAPDH вперед (GAA GGT GGT GAA GCA GGC AT), GAPDH-обратно (GCA TCG AAG GTG GAA GAG ГТГ). Реакция объема (25 мкл) был сделан с 2 ? QuantiTect SYBR Green ® Master Mix (Qiagen, Hilden, Германия, http://). Y-хромосома и GAPDH стандарт с известной концентрацией ДНК были включены в анализ. Отрицательный контроль - женская ДНК и шаблон были также включены. Реакции: денатурация (95 ° C в течение 15 минут), циклы (40 повторов, первый шаг при 95 ° C в течение 20 секунд, второй шаг при 56 ° С в течение 20 секунд, а третьему шагу при 72 ° C в течение 30 секунд приобретение SYBR зеленый сигнал), стабилизация (60 ° C в течение 1 минуты), расплавление (от 60 ° C до 95 ° C 5 секунд на первый шаг, 5 секунд на следующих шагах)
Результаты.
Селективное выживание и обогащение MGMT (P140K) преобразуют мышечные клетки с селекцией BCNU и O6BG In Vitro
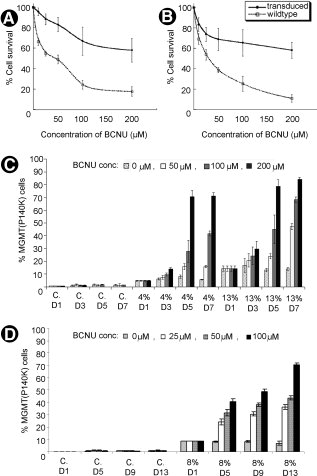
Мы сначала пытались определить может ли принудительная экспрессия MGMT (P140K) в C2C12 клетках и миобластах человека обеспечивать защиту от BCNU в результате селективного обогащения преобразованных клеток. Когда трансдуцированные и нетрансдуцированные клетки обрабатывали различными дозами BCNU (0 мкм, 50 мкм, 100 мкм, 200 мкм) и O6BG (40 мкм), анализ пролиферации клеток показал, что C2C12 (рис. 1А) и человеческие первичные миобласты (рис. 1В), экспрессирующие MGMT (P140K) показали большую устойчивость к BCNU и O6BG по сравнению с клетками дикого типа. Чтобы проверить селективное обогащение MGMT (P140K)-экспрессирующих клеток, C2C12 культур, содержащих 0, 4, или 13% MGMT (P140K) преобразованных клеток на фоне клетки дикого типа, клетки были обработаны четырьмя концентрация BCNU в течение 7 - дневного периода (рис. 1в). Аналогично, человеческие первичные культуры миобластов содержащих 0 или 8% MGMT (P140K)-трансдуцированных клеток обрабатывали тремя концентрациями BCNU за 13-дневный период (рис. 1D). FACS анализ показал выборочное обогащение MGMT (P140K)-преобразованных клеток с течением времени в обеих культурах, которые коррелируют с количеством MGMT (P140K)-преобразованных клеток изначально присутствующих и дозами BCNU. Для обоих комплектов культур, более высокие дозы BCNU достигнули лучшего выбора, чем при более низких дозах. Эти результаты также подтверждают, что EF-1? промоутер экспрессии MGMT (P140K) активен, как в мышиных, так и в человеческих миогенных клетках.
Рисунок 1
![]()
На дифференцировку миобластов не влияет последующая ретровирусная трансдукция и BCNU/O6BG лечение
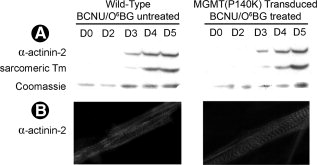
Чтобы определить можети ли ретровирусная трансдукция MGMT (P140K), а также BCNU/O6BG лечение миобластов поставить под угрозу их способность к дифференциации, MGMT (P140K)-трансдуцированные C2C12 клетки обрабатывали BCNU/O6BG, что не помешало им дифференцироваться в мышечные трубки. Потенциал дифференциации MGMT (P140K)-преобразованных клеток был сравненим с необработанным диким типом C2C12 клеток. Дифференцированные клетки были проанализированы для выявления экспрессии специфических белков мышц тонкой нити, ?-актинина-2 и саркомерного тропомиозина. Уровни экспрессии для ?-актинина-2 и саркомерного тропомиозина были сопоставимы между трансдуцированными ( BCNU/O6BG лечение) и диким типом ( BCNU/O6BG необработанные) клеток после начала дифференциации (рис. 2), что свидетельствует о скорости и степени изменения дифференциации. Кроме того, иммунохимическое окрашивание ?-актинина-2 показало нормальное расположение саркомеров в MGMT (P140K) трансдуцированных клетках, обработанных BCNU/O6BG (рис. 2В). BCNU/O6BG лечение дикого типа C2C12 культур отображает существенную гибель клеток и с нарушением последующей дифференциации(данные не показаны).
Рисунок 2
Затруднения регенерации дикого типа мышц, обработанных BCNU + O6BG, в то время как MGMT (P140K)-экспрессирующие мышцы сохранялись.
BCNU алкилирует ДНК внутри клетки независимо от состояния клеточного цикла, однако, результаты ДНК алкилирования проявляются только тогда, когда клетка подвергается пролиферации. Скелетные мышцы находятся в состоянии покоя в нормальных условиях, но становятся митотически активны во время регенерации. Чтобы
![]()
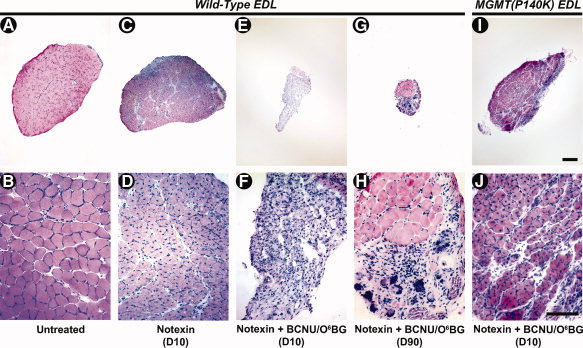
получить доказательства правильности принципа того, что MGMT (P140K) может защитить от BCNU алкилирования у животных, мы использовали трансгенные линии мышей, у которых MGMT (P140K) экспрессируется во всех типах клеток. Мы индуцировали регенерацию в EDL мышцах дикого типа и MGMT
![]()
(P140K) тг / тг мышей, введенный им инъекций нотексин агента (0,1 мкг) с и без BCNU + O6BG. После 10 дней регенерации в случае отсутствия препарата, гистологические сечения дикого типа мышц показывали регенерирующие мышечные волокна с внутренними ядерами, отличительной чертой нормальной регенерации мышечных волокон (ср. рис. 3C, 3D с 3А, 3Б). Когда дикому типу EDL вводили нотексин и BCNU (0,2 мг, IM) + O6BG (30 мг / кг б. Мас., IP), мышцы показали полное отсутствие регенерации мышечных волокон до 10 дней после обработки (Рис. 3E, 3F). Эта неспособность к регенерации после лечения сохранялась через 90 дней (рис. 3G, 3H). Это свидетельствует о том, что удаление спутниковых клеток BCNU/O6BG приводит к постоянной неспособности к регенерации мышц. В противоположность этому, когда же процедура была применена к EDL мышцам MGMT (P140K) трансгенных мышей, регенерирующие мышечные волокна явно присутствовали 10 дней после лечения (рис. 3I, 3J). Хотя степень восстановления в среде обработанных MGMT (P140K) мышц трансгенных мышей была уменьшена по сравнению с диким типом мышц, регенерирующим за тот же промежуток времени в отсутствие препаратов (ср. рис. 3D с 3J), эти результаты подтверждают достаточное выражение MGMT (P140K) в клетках, которые способствуют регенерации мышц
Рисунок 3
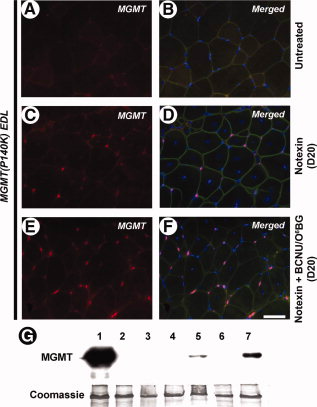
Иммунное обнаружение человеских MGMT выявило ряд MGMT (P140K)-экспрессирующих клеток в регенерации мышц MGMT (P140K) трансгенных мышей (рис. 4C, 4D) по сравнению с необработанными мышцами (рис. 4А, 4В). Поскольку экспрессия MGMT (P140K) трансгенов обусловлена ??EF-1? промоутером, который действует только в митотически активных клетках, экспрессия MGMT (P140K) отсутствует в необработанных мышцах. После BCNU/O6BG лечения, регенерирующие MGMT (P140K) трансгенные мышцы имеет значительно большее количество MGMT-экспрессирующих клеток (рис. 4E, 4F) с указанием наувеличениепопуляции этих клеток при выборе давления, оказываемого BCNU/O6BG лечением. Вестерн-блот анализ подтвердил увеличение количества MGMT белка, присутствующего в MGMT (P140K) трансгенных EDL обработанных нотексином и BCNU/O6BG (рис. 4G, переулок 7) по сравнению с обработанными только нотексином (рис. 4G, Lane 5). Необработанные MGMT (P140K) трансгенные мышцы дикого типа, и мышцы, обработанныенотексином с или без BCNU/O6BG не содержат обнаруживаемых уровней MGMT (рис. 4G, дорожки 2, 3, 4, 6).
Рисунок 4
![]()
Выделение и характеристика CD34 донорских клеток из регенирирующих скелетных мышц
CD34 антиген клеточной поверхности используется для выбора донорских клеток, так как многие недавно идентифицированные мышечные стволовые клетки обычно экспрессируют этот поверхностный антиген. Этот выбор стратегии позволил отобрать максимально возможное число донорских клеток в естественных условиях с использованием одного маркера, сведя к минимуму время необходимое для сортировки, при максимальной жизнеспособности клеток. Число клеток CD34,которые могут быть изолированы из нормальных, неповрежденных мышц было ниже 1 ? 105 клеток в мышцах TA (рис. 5). Регинереирующая ТА взятаячерез 3 дня после инъекции нотексина, содержала на порядок больше клеток CD34, примерно 11 ? 105 клеток в мышцах TA (рис. 5). В рамках этого увеличения популяции клеток CD34, две различные субпопуляции стали различимы после ночной культивации. Одна популяция - прилипшие клетки отображают фибробластоподобную морфологию (рис. 5а), в то время как вторая популяции маленькие, круглые, преломляющие свет клетки, неадгезивные (рис. 5б). Вестерн-блот анализ показал, высокую экспрессию MyoD, маркера миогенной клетки (рис. 5). Кроме того, прилипшие клетки экспрессировали определяемые уровни миогенина, маркера терминальной дифференцировки мышц (рис. 5).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
