
Рисунок 5
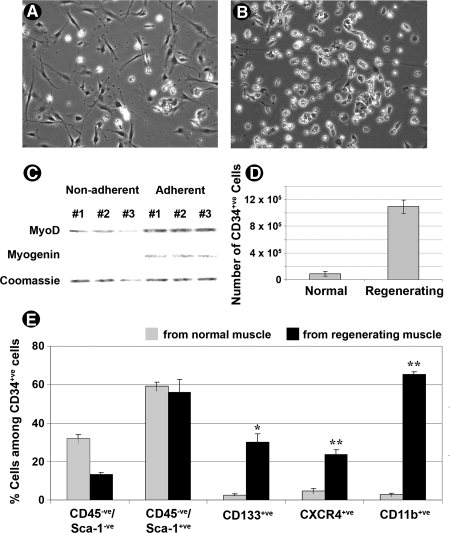
Мы также характеризовали CD34 + популяцию с использованием FACS анализов. В ряде исследований сообщается, что полученные стволовые и спутниковые предшественники мышечных клеток у мышей обычно выражают CD34 и не экспрессируют CD45. Среди них субпопуляции спутниковых клеток предшественников не экспрессируют Sca-1, в то время как другие, такие как мышечные стволовые клетки, мезоангиобласты все экспрессируют Sca-1. FACS анализ показал, что CD45-ve/Sca-1-ve клетки составляли примерно 13% CD34 + VE клеток, выделенных из мышц регенерирующим по сравнению с 31% в неповрежденных мышцах (Рис. 5E).Представление CD45-ve/Sca-1 + субпопуляции было одинаково в неповрежденных и регенерирующих мышцах у 59 и 56%, соответственно (Рис. 5E).
Немышечные клетки, таких как эндотелиальные предшественники, моноциты и макрофаги привлекаются к месту травмы в скелетных мышцах. Эти клетки играют важную роль не только в развитии активации и пролиферации стволовых клеток, а также в удалении ткани мусора и участии в реваскуляризации. Для оценки представления таких опорных клеток в CD34 + популяции FACS анализ проводился для количественной оценки доли CD133 + е (маркер эндотелиальных клеток-предшественников), CXCR4 + е (рецепторы для цитокинов стромальные, полученные фактор-1 [SDF -1]) и CD11b + VE (маркер для моноцитов). Каждая из этих трех подгрупп составила очень небольшую часть CD34 + VE клеток, выделенных из неповрежденных мышц примерно на 3% -5% (Рис. 5E). Тем не
![]()
менее, при мышечной регенерации процент значительно увеличится в CD34 + популяции: 30% CD133 + , 24% CXCR4 + , 66% CD11b + клеток (Рис. 5E). Наличие этих опорных клеток в значительных количествах в CD34 + популяции клеток из регенерирующих мышц может оказаться полезным для распространения донорских стволовых клеток после трансплантации.
Увеличенное выживание CD34 + MGMT (P140K) донорских клеток и образование De Novo мышечных волокон
![]()
Чтобы проверить, может ли защита и селективное обогащение MGMT (P140K)-экспрессирующих клеток происходить в естественных условиях после трансплантации, доноры CD34 + клеток из мышцы MGMT (P140K) тг / тг или C57BL/6JArc мышей дикого типа пересаживают в EDL мышцы реципиента дикого типа C57BL/6JArc мышей. Донорские клетки вводятся в мышцы реципиента вместе с нотексином (0,2 мкг), чтобы вызвать дегенерацию здоровых мышц. Мыши реципиенты были разделены на три группы с пятью мышами или более в каждой группе: Группа А (дикий тип клеток донора только), Группа B (дикий тип донорских клеток с BCNU + O6BG), Группа C (MGMT [P140K] тг / тг клеток донора с BCNU + O6BG).
Мышцы реципиента были собраны через 7 дней после трансплантации, а также процент мужской ДНК был определен с использованием количественной ПЦР. В группе А (n = 5), небольшая часть от общего числа ДНК (0,39% ± 0,05% SD) был донорского происхождения (рис. 6, группа А). Эта группа представляет собой обычный протокол трансплантации донорских клеток, которые пересаживают в мышцу получателя без какого-либо отбора. В группе В (n = 8), высокая доля ДНК донора (2,1% ± 0,6% SD), возможно, отражает цитотоксический эффект BCNU/O6BG лечения на мышцы регенерирующей эндогенной популяции стволовых клеток (рис. 6, Группа B). В этой группе только донорские клетки, которые вышли из клеточного цикла во время трансплантации выжили бы, так как пролиферирующие клетки дикого типа были также подвержены цитотоксическому эффекту BCNU/O6BG во время пересадки. Мышцы реципиентов из группы С (п = 10) содержат самый высокий процент (6,4% ± 1,7% SD) донорской ДНК (рис. 6, группа C), что свидетельствует о защитном эффекте MGMT (P140K) против цитотоксических эффектов BCNU / O6BG, и привело к селективному выживанию и обогащению донорских клеток
Рисунок 6
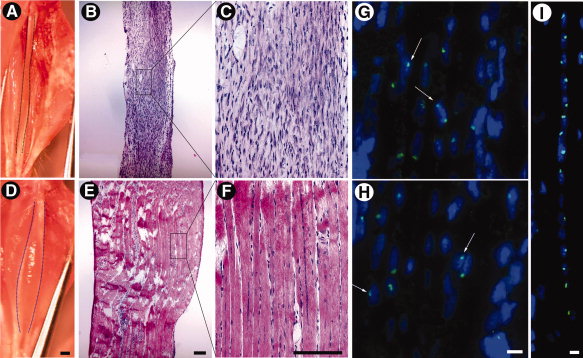
Когда мышцы из группы B (n = 5) и С (п = 5) были собраны через 14 дней после трансплантации, мышцы из двух групп показали замечательную разницу в размерах (ср. рис. 7А и 7D). Гистологическое исследование показало отсутствие нормальной реакции восстановления в мышцах группы B (дикий тип доноров), большинство ткани занимают мононуклеарные клетки (рис. 7б, 7в). Это говорит о том, что повышение количества клеток дикого типа донора в BCNU/O6BG-обработанных мышцах не проявляется как повышение регенерации мышц. В отличие от группы C мышц с донорскими клетками MGMT (P140K) трансгенных мышей, показавших нормальную реакцию регенерации, с обильными мышечными волокнами, содержащими централизованно ориентированные ядра, что свидетельствует о регенерации волокон (рис. 7E, 7F).
Рисунок 7
Дискуссия.
MGMT (P140K)-опосредованное обогащение клеток доноров после трансплантации стволовых клеток в скелетные мышцы
Наши результаты дают доказательство правильности концепции того, что MGMT (P140K)-опосредованная BCNU/O6BG селективная стратегия обогащения может быть успешно применена в соматических стволовых клетках скелетных мышц. Эта стратегия была разработана для прижизненного обогащения гемопоэтических стволовых клеток с доказанной эффективностью. Негемопоэтическое применение стратегии на сегодняшний день показало, что клетки костного мозга экспрессирующие MGMT (P140K) можно прививать в легких. Здесь мы покажем, успешное приживление MGMT (P140K) экспрессирующих CD34 + клеток после трансплантации скелетных мышцах, обрабатанных алкилирующим агентом. Насаждение популяции клеток донора может включать в себя клетки спутника, а также другие стволовые клетки, экспресирующие антиген CD34 на момент изоляции. Кроме того, другие немышечные типы клеток в пределах CD34 + популяции клеток-доноров могли бы обеспечить косвенную поддержку приживления донорских мышечных стволовых клеток и образование волокон.
В предыдущих исследованиях с участием трансплантации стволовых клеток, выживание клеток донора в трансплантации хозяину было одной из основных проблем. Выживание и приживление MGMT (P140K)-экспрессирующих донорских клеток, показаное в этом исследовании, подтверждает важность восприимчивости хозяина в начале выживания и последующего распространения донорских стволовых клеток. Концепция кондиционирования принимающей ниши привела к использованию нелетальных доз облучения для абляции деятельности клеток-хозяина. Тем не менее существуют два основных аргумента против такого метода:в том числе вопрос безопасности применения высоких доз облучения для достижения значимого кондиционирования ниши хозяина и недискриминационный характер облучения, в результате чего клетки донора в равной степени подвержены повреждениям, если последующие дозы облучения применяются. Эта стратегия может обеспечить эффективную терапию для лечения заболеваний, при которых конкретные мышцы страдают, таких как окуло-глоточная мышечная дистрофия, в сочетании с вирусной трансдукцией донорских стволовых клеток для исправления генетических мутаций. Кроме того, ограниченная локальная доставка алкилирующих агентов, используемыех в этом исследовании, подчеркивает безопасность этого способа доставки. Это обнадеживает, учитывая, что клеточная терапия для замены неблагополучных взрослых стволовых клеток вследствие старения и в результате различных генетических / дегенеративных заболеваний в настоящее время рассматривается во многих различных типах тканей, включая легкие расстройства, хронические травмы печени, желудочно-кишечные расстройства и I и II тип сахарного диабета. Наши данные подтверждают возможность применения MGMT-опосредованной стратегии обогощения клеток донора в других типах тканей и органов, где клеточная терапия существует как возможный вариант лечения.
![]()
Индукция эндогенной регенерации мышц после трансплантации донорских клеток CD34 + экспрессирующих MGMT (P140K)
Неожиданное открытие, что эндогенные клетки вносят значительный вклад в регенерацию BCNU/O6BG-обработанных мышц получателя после трансплантации MGMT (P140K)-экспрессирующих клеток донора возникает вопрос, какие факторы и / или типы клеток маршалируются MGMT (P140K)экспрессирующими клетками и почему? В отсутствие пересаженных MGMT (P140K)-экспрессирующих клеток, эта мышца не в состоянии самостоятельно регенерировать после травмы и медикаментозного лечения (за этим последовало до 90 дней, на рис. 3G, 3H). Мы полагаем, что фактор (ы), которые выделяется при образовании донорно-производных мышечных волокон, привлекают клетки, которые способны к миогенной дифференциации. Одним из таких кандидатов является ?-хемокин (SDF-1), хемоаттрактант для клеток с G-белками рецептора CXCR4. Это хорошо установлено, что SDF-1 освобождается из регенирирующих мышц, но что более важно, что недифференцированные спутниковые клетки мышц экспрессируют CXCR4 и реакция хемоаттрактанта обеспечивается наличием SDF-1 через фосфорилирование p42 / 44 митоген-активированной протеинкиназы (МАРК) и AKT серин-треонин киназы.
Поэтому вполне возможно, что клетки-сателлиты, проживающих в мышцах, прилегающих к EDL получают MGMT (P140K) донорских клеток мигрируют в эту мышцу и способствуют регенерации в ответ на освобождение SDF-1 из волокон, образованных из донорских клеток. Тем не менее, количество этих клеток, проживающих в соседних мышцах будет сокращено, потому что при местных внутривенных инъекциях с последующей 2-х минутной блокадой кровотока в проксимальном отделе задних конечностей были бы подвержены все соседние группы мышц (и их резидентные клетки спутники) в задней конечности последствиям BCNU. После алкилирования клетки не смогли бы пройти деление, тем самым уменьшая вклад сателлитных клеток из соседних мышц.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
