


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Первичная роль функциональной ишемии, количественные данные для двух механизмов развития и терапии ингибиторами фосфодиэстеразы 5 мышинной модели мышечной дистрофии.
Введение
Мышечная дистрофия Дюшенна (МДД ) обусловлена ??отсутствием продукта гена дистрофина, и затрагивает приблизительно одного из 3500 новорожденных мальчиков. В скелетных мышцах МДД пациентов проходит медленное прогрессивное повреждение, которое приводит к возникновению заболевания. Точная патофизиологии не известна, кроме широко принятой теории, что основную роль играет мембранная уязвимость. Предшествующие исследования показали, что отсутствие дистрофина и связанных с ним молекул вызывает дефекты в потое крови в мышечные ткани. В ответ на сократительную нагрузку нормальные мышцы стремятся к увеличению притока крови для удовлетворения метаболических потребностей. Однако, когда этот ответ ослабляется, мышцы имеют риск ишемии из-за отсутствия достаточного количества кислорода и питательных веществ или достаточного выведения накопленных метаболитов, патологическое состояние "функциональной ишемии ». Ишемия определяется как снижение кровотока из-за структурных обструкций сосудов или вазоконстрикции. Состояние функциональной ишемии - когда приток крови не может сравниться с метаболическим спросом тканей даже при отсутствии обструкции сосудов. Баланс между спросом и предложением кровотока нарушается в обоих случаях. Оксид азота (NO) , также известный как эндотелий - расслабляющий фактор ( ЭФР ) , производимый в скелетных мышцах контролирует местный кровоток в мышцах [8] , [9] наряду с различными другими вазорегуляторными молекулами. У пациентов с МДД, а также мышей MDX ( мышиный эквивалент ), экспрессия сарколеммой нейрональной синтазы оксида азота ( нОАС ) в скелетных мышцах значительно снижается с сопутствующими нарушениями в регулировании кровотока. Различные исследования показали, что имеют место сосудистая патология, изменения сосудорасширяющего ответа и нарушенная сосудорасширяющая сигнализация нОАС. Не было оценено в деталях, однако, является ли нарушение регуляции кровотока в связи с отсутствием экспрессии ННО основной причиной или вторичным дефектом мышечной дистрофии. У мышей с отсутствием ННО схожие аномалии кровотока не проявляются фенотипом мышечной дистрофии, было высказано предположение, что функциональная ишемия или отсутствие ННО может быть вспомогательным мероприятием, но не является прямой причиной заболевания. Отсутствие фенотипа дистрофии у мышей с отсутствием фермента, однако, означает не более, чем отсутствие ННО или недостаточность кровотока, чтобы вызвать мышечную дистрофию. Было бы неправильно сделать вывод, что отсутствие ННО или нарушение кровообращения не является существенной причиной заболевания. В дополнение к сказанному, ткани продуцируют другие типы сосудорасширяющего факторов, включая эндотелиальный фактор гиперполяризации ( EDHF ). Хотя структура EDHF остается неясной, предыдущие доклады показали, что супероксиддисмутаза (СОД ) производит перекись водорода (H2O2) , которая оказывает EDHF - подобные функции. Кроме того, было предложено, что одного фактора недостаточно для объяснения патогенеза МДД и предположили, что, по крайней мере, два фактора, необходимы для побуждения повреждения мышечных волокон. В этом исследовании мы оценили регулирование потока крови в патогенезе мышечной дистрофии с использованием естественных микроскопических анализов : мы исследовали, как кровоток реагирует на сокращение мышцы у MDX и контрольных мышей, с NO/H2O2 производством в мышцах, ослабляется ли у MDX мышей, и является ли увеличение окиси азота фактором, который может предотвратить функциональную ишемию и повреждение мышечных волокон у MDX мышей. Степень функциональной ишемии и количество повреждений миофибрилл сравнивались у MDX и контрольных мышей.
Результаты
Контрольные мыши показывают увеличение потока крови в мышцах при сокращении, чего нет у MDX мышей.
Различные параметры существуют для анализа местного кровотока, в том числе поток эритроцитов, скорость потока плазмы, диаметр кровеносного сосуда и функциональная плотность капилляров. Среди них потокэритроцитов отражает местную подачу кислорода и является хорошо организованным стандартным методом для оценки локального кровотока. В этом исследовании мы измерили периферийный поток эритроцитов в внутримышечных артериолах в естественных условиях ( номенклатура кровеносных сосудов, см. дополнительную информацию, S1 рисунок) . Наблюдаемые люминесцентные эритроциты, помечены различными красителями включая PKH26GL, что является общепринятым методом анализа микроциркуляции. Чтобы оценить возможное влияние флуоресцентных меченных эритроцитов PKH26GL на динамику потока эритроцитов, альтернативный способ окрашивания ( FITC) был использован по той же схеме кинетики (данные не показаны). Базальный уровень абсолютного потока ( окрашенные и не окрашенные эритроциты комбинированный) был 8043,2 ± 1372,5 и 10403,2 ± 1876,0 для контроля и MDX мышей ( потока значение в минуту ± SE, N ( количество животных ) = 18 и 17 , статистически не отличались, р = 0,31 при т - тест) . Справедливость нашего метода маркировки, таким образом, обеспечивает согласованность с предыдущими исследованиями потока эритроцитов для венулы ( 12-39 мкм в диаметре ) прибл 100000 в минуту у мышей [ 25], опираясь на капиллярный поток 1800 или 1200-2000 в минуту у крыс [26] или хомяков [27] (заметим, что одна первичная артериола питает несколько капилляров и множество капилляров впадают во вторичные венулы). Как показано на рисунке 1а, у контрольных мышей произошло увеличение потока, что продолжается более 8 - 10 минут в связи с сокращением в ответ на 50 Гц стимуляцию. Там не было никаких существенных различий в структуре изменения потока между NMJ и не NMJ районах либо штаммами мышей, за исключением небольшой разницы во времени увеличения потока у контрольных мышей. Таким образом, все измерения проводились в не - NMJ областей, если не указано иное. В предыдущих экспериментах, высокие частоты стимуляции при 20 Гц на мышцы задней конечности крыс привело к увеличению скорости кровотока продолжительностью до 14 минут после прекращения стимуляции [28].
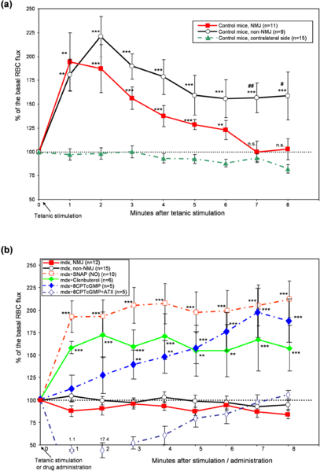
Прямая стимуляция на одной стороне грудино-сосцевидной мышцы не вызывала сокращение на другой стороне, а поток в контралатеральной стороне остался на базальном уровене (рис. 1а). Это наблюдение позволяет предположить, что в условиях в этом исследовании, стимуляция грудино-сосцевидной мышцы не изменяет сердечный выброс, и увеличение кровотока в соответствии с местной стимуляцией мышц. Эксперименты показали, у MDX мыши полное отсутствие увеличения потока после стимуляции как в NMJ, так и в не NMJ областях (рис. 1b ), несмотря на тот факт, что тетанические стимулы дали такой же степени сокращения мышц у контрольных мышей (рис. S2 , вспомогательная информация ). Изучение вопроса отсутствия реакции у MDX мышей было связано с дефектом кровеносных сосудов или дефектом в передаче сигнала между мышцами и кровеносными сосудами, мы применили SNAP (S - нитрозо - N - ацетилпенициламин, NO донор) на местном уровне. Когда SNAP был применен в покое на мышцы MDX мышей, поток был увеличен до максимума 212,4 % от базального потока (рис. 1b). Это увеличение было сопоставимо с изменениями у контрольных мышей, которым вводили такую ??же дозу SNAP (данные не показаны, максимальное увеличение до 236,2 %, N ( количество животных ) = 10, существенно не отличались от MDX мышей в любой момент времени между 0 и 8 минут). Этот результат позволяет предположить, что сосудорасширяющие механизмы в кровеносных сосудах были функциональны у MDX мышей, но сигналы между скелетной мышцей и кровеносными сосудами были скомпрометированы. Дальнейшее подтверждение этого вывода наблюдалось, когда мембранпроницаемый цГМФ аналог, 8-( 4- Хлорфенилтио ) - гуанозин-3 ', 5'- циклического монофосфат (8- СРТ цГМФ) был локально применен. Этот препарат вызвал медленное увеличение потока у мышей MDX, достигнув такой же степени реакции, которая наблюдалось в группе SNAP через 7 минут после введения. Учитывая, что сосудорасширяющий
![]()
эффект NO через цГМФ - зависимые пути [ 29] - [ 31] , различие в кинетике увеличения потока между группами с SNAP и с 8 - CPT цГМФ произошло, вероятно, из-за разницы в скорости достижения препаратами своих клеток-мишеней. Влияние на увеличение потока эритроцитов 8 - CPT цГМФ неспецифически необратимо сосудосуживающее действие, но, скорее всего, определенно физиологическим регулированием, потому что этот ответ полностью противодействует дальнейшему добавлению физиологической концентрации вазоконстриктора ангиотензина - II ( ATII ) . Поскольку ?2 - агонисты имеют различный механизм вазодилатации, мы рассмотрели кленбутерол, как препарат, который вызывает расширение кровеносных сосудов у MDX мышей. Когда кленбутерол был локально введен вместо SNAP, MDX мыши показали увеличение потока, хотя и в меньшей степени ( максимум до 172% , рис 1b). В результате этот эксперимент подтверждает, что механизм вазодилатации гладких мышц сосудов является функциональным у MDX мышей.
Оксида азота и производство перекиси водорода в ответ на сокращение мышц ослабляется у MDX мышей.
В естественных условиях производство сосудорасширяющих молекул (т. е. нет и H2O2 ) измерялось в грудино-сосцевидной мышце MDX и контрольных мышей для исследования механизмов функциональной ишемии у мышей MDX. Прямая тетаническая стимуляция грудино-сосцевидной мышцы индуцировала заметное повышение уровня азота в мышечных волокнах (рис. 2- б). Когда L - NAME ( нОм - нитро - L - аргинина, метиловый эфир ) ,неспецифический ингибитор NOS, был применен, производство NO в мышечных волокнах было снижено примерно на 67,6% из чистого прироста. Тот факт, что экзогенный L-NAME не может подавить все внутриклеточное NO производство, согласуются с предыдущими исследованиями [33]. Последствия сокращения мышц между контролем и MDX мышами (рис. 2и B). Базальный уровень NO в покоящейся мышце MDX мышей был выше, чем у контрольных мышей, но увеличение NO производства после сокращения мышц было полностью прекращено у MDX мышей. Производство NO от не мышечными клетками было отмечено ( черно-белая стрела на рисунке 2а).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
