
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Иммуноокрашивание и обнаружение центральных ядер.
EDL сечения (10 мкм) с контрольной группы и получавшей АЦ MDX мышей и подобранных по возрасту мышей дикого типа были использованы как для иммунного окрашивания дистрофина, и визуализации центральных ядер. Подробности процедуры были описаны ранее (Yeung соавт. 2005). Во-первых, поперечные сечения инкубировали
![]()
с AffiniPure Fab и Fc-фрагментами (Jackson ImmunoResearch лаборатории, West Grove, PA, США) в концентрации 60 мкг л в 2% БСА в течение 30 мин, затем промывали в фосфатно-солевом буфере (ФСБ ), а затем блокировали в
![]()
2% БСА в течение 15 мин. Ранее нами было показано, что эта процедура предотвращает неспецифическое связывание антимышиными вторичными антителами к эндогенным иммуноглобулинам мыши в срезе ткани. В качестве первичных антител были использованы мышиные моноклональные антитела против: кавеолина-3, разбавленного 1: 50 (BD Transduction Laboratories, США), ?-дистрогликана, разбавленного 1: 20 (NCL-би-DG, Novocastra лаборатории, Ньюкасл, Великобритания) и атрофина, разбавленого 1: 20 (NCL-Drp2, Novocastra Laboratories). Вторичные антитела были козьими антимышиными конъюгированными с Су3 антителами, разбавленными 1: 200 (Jackson ImmunoResearch Laboratories). Для обнаружения ядер вторичные антитела содержали флуоресцентный окрашивающий ядра 4 ',6-диамидино-2-фенилиндол (DAPI, Sigma, Сент-Луис, штат Миссури, США), который был разбавлен 1: 100. Флуоресцентные изображения поперечных сечений были приняты с 5 ? или 20 ? (Zeiss плана Neola), и количество мышечных волокон с центральными ядрами рассчитывали как процент от общего количества волокон с видимыми ядрами (т. е. оба центральных и периферических).
Окрашивание дигидроетидиумом на АФК.
Дигидроетидиум (ДГЭ) является широко используемым показателем продукции АФК как в пробирке (Benov соавт. 1998), так и в естественных условиях (Robinson и соавт. 2006). Он окисляется АФК, в частности супероксидом (О2-), образуя бромид этидия, который флуоресцирует красным (Benov соавт. 1998). Поэтому интенсивность флуоресценции в ядрах клеток является мерой продукции АФК. АФК измеряли путем инкубации EDL сечений с 5 мкм ДГЭ в PBS при 37 ° С в течение 30 мин. ДГЭ интенсивность оценивали путем подсчета количества пикселей превышающий заданный порог, который был создан с целью устранения помех от любой фоновой флуоресценции.
Вестерн-блоттинг.
Замороженные мышцы TA лизировались Polytron PT 1200 гомогенизатором (Kinematica, Littau / Люцерн, Швейцария) с использованием ледяного буфера для лизиса либо общего лизата или ядерной фракции. Для полного лизата мышцы лизировали в буфере, содержащем 50 мМ Трис, рН 7,5, 150 мм NaCl, 25 мМ ЭДТА, 25 мм EGTA, 1% Тритон Х-100, ингибитора протеазы, калпаин, ингибитора фосфатазы (Sigma) . Через 30 мин инкубации гомогенаты центрифугировали при 15 800 g в течение 30 мин при 4 ° С, и супернатант удаляли. Для ядерных фракций два буфера были использованы. Во-первых, мышцы лизировали в буфере, содержащем 10 мМ Hepes, 1,5 мМ MgCl2, 10 мМ KCl, 0,5 мМ DTT, 0,05% NP40, протеазы, калпаина 1 и фосфатазного ингибитора. Мышечные лизаты хранили на льду в течение 10 мин и затем центрифугировали при 800g в течение 10 мин при 4 ° C. Надосадочную жидкость удаляли, а осадок, содержащий ядерные белки, затем ресуспендировали в буфер, используемый для общего лизата. Через 30 мин гомогенаты центрифугировали при 15 800 g в течение 30 мин при 4 ° С и супернатант удаляли. Концентрацию белка в надосадочной жидкости определяли с помощью анализа Брэдфорда (Bio-Rad, Hercules, CA, USA).В общей сложности 20 мкг белка на лунку загружали в 4% (укладки) -12% (решить) полиакриламидном геле. Белки разделяли электрофорезом в ДСН-ПААГ и переносили на нитроцеллюлозную мембрану с использованием мини Транс-блот передачи (Bio-Rad). Мембраны блокировали в течение 30 мин при 5% обезжиренного сухого молока в PBS-Tween 20 при комнатной температуре. Для полного лизата мембраны инкубировали с первичными антителами против кавеолина-3, разводили 1: 1000 в блокирующем буфере в течение 1 ч при комнатной температуре. Ядерные фракции инкубировали с первичными антителами к NF-kB (p65 фосфо S276, Abcam, Cambridge, UK), разбавленными 1: 500 в блокирующем буфере в течение ночи при 4 ° С. После промывки PBS-T, мембраны были инкубированы в течение 1 ч при комнатной температуре с HRP-конъюгированными анти-мышинными (для кавеолином-3) или анти-кроличьими (для NF-kB) вторичными антителами (Santa Cruz Biotechnology, Santa Cruz, CA, USA) в разведении 1: 1000. Мышиные моноклональные ?-актиновые антитела (Sigma) использовали в качестве контроля для обеспечения равной нагрузки.
Статистика
Все данные представлены как среднее значение ± S. E.M. Значительная разница между группами была определена с использованием критерия Стьюдента для непарных данных или двухфакторного дисперсионного анализа. Уровень значимости был установлен на уровне р <0,05.
Результаты
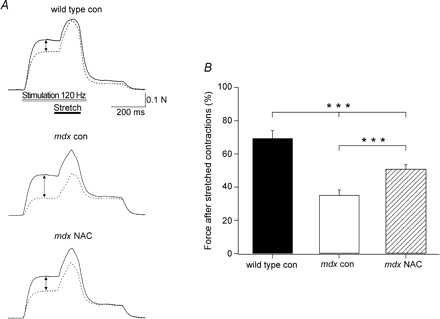
MDX мышцы обработанные АЦ улучшают силу после сокращения.
Сокращения проводили при 35 ° С, а не при комнатной температуре, так как АФК в изолированных скелетных мышцах, как известно, значительно возрастет в этом диапазоне температур (Arbogast и Reid, 2004). В предварительных экспериментах, мы обнаружили, что снижение силы мышц MDX от трех растянутых сокращений при 35 ° С было сопоставимо (~ 35%). Поэтому мы решили использовать три сокращения в нынешней серии экспериментов. Как показано на рис. 1А, сила сокращения вызванного растяжением было значительно больше в MDX мышцах, чем дикий тип. Через 60 мин после сокращения (фиг. 1В), среднее усилие было 35 ± 3% для мышц MDX контроля (n = 9), который был значительно меньше, чем у MDX мышц с АЦ, где величина составила 51 ± 2% (Р <0,001, п = 11). Дикий тип мышц имел среднюю силу 69 ± 4% (n = 4), значительно больше, чем оба MDX (Р <0,001). Для того, чтобы установить, что АЦ-индуцированная защита от стресс-индуцированной потери силы была присуща MDX мышцам, мы также провели несколько экспериментов с АЦ у дикого типа мышц. АЦ не оказал существенного влияния на снижение силы мышей дикого типа (72 ± 1%, N = 6).
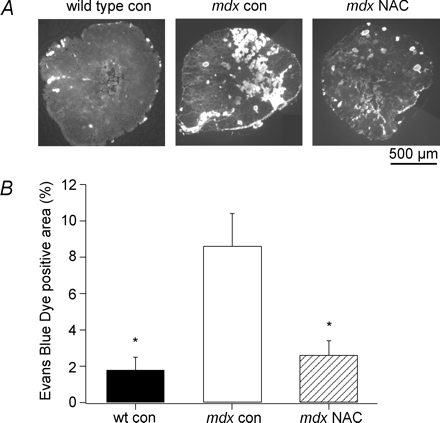
АЦ предотвращает поглощение СКЭ в MDX мышцах после сокращения.
Поступление в мышечные волокна СКЭ было использовано для оценки повышенной проницаемости мембраны у MDX и дикого типа мышц после сокращения. EDL мышцы были заморожены через 60 мин после растяжения, и сечений, взятые из середины мышцы живота, были использованы для количественного анализа СКЭ. На рис. 2А, сечения показывают, что MDX контрольные мышцы имели значительно больше СКЭ-положительных волокон, чем дикий тип, в то время АЦ предотвращает увеличение поглощения красителя в MDX мышцах. Когда данные были собраны (фиг.2В), мышцы MDX контроля имели 8,6 ± 1,8% (N = 8) СКЭ положительных волокон по сравнению с 1,8 ± 0,7% дикого типа мышц (п = 5, P <0,05) и до 2,6 ± 0,8% для MDX мышц АЦ (N = 7, р <0,05). Эти данные предполагают, что АФК способствуют наиболее повышенной проницаемости участка мембраны в MDX мышц при 35 ° С.
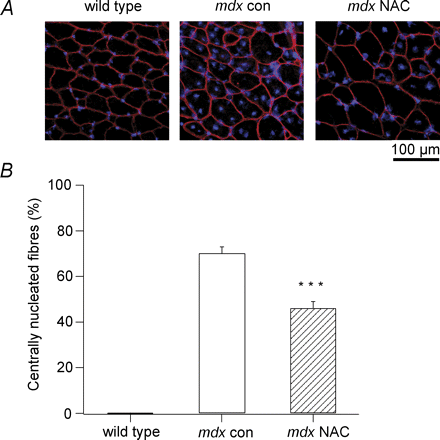
Лечение АЦ снижает содержание центральных ядер у MDX мышей
Регенерацию мышечных волокон после периода некроза можно количественно определить по наличию центральных ядер в сечениях мышц. В этих экспериментах MDX мышей обрабатывали 1% АЦ в питьевой воде от 3 до 9 недель - в период наиболее обширного повреждения и регенерации. В сечении мышц от MDX мышей в возрасте 9 недель было большое количество волокон, содержащих центральные ядра, и заметно меньше в MDX мышцах мышей, получавших АЦ (фиг.3А). Среднее число волокон с центральными ядрами составляло 70 ± 3%
![]()
для MDX мышц контроля (N = 6), и было значительно сокращено до 46 ± 3% в MDX получавших АЦ (N = 6, р <0,001, рис. 3В). Учитывая, что дикий тип мышц не имел центральных ядер (рис. 3), этот результат показывает, что в мышцах MDX мышей, получавших АЦ, было примерно на одну треть меньше поврежденных волокон, чем у MDX мышей контроля.
Лечение АЦ защищает от потери сил при сокращениях.
В эксперементах на изолированных мышцах (как описано ранее) АЦ обеспечивает защиту против снижения силы MDX мышц. Таким образом, мы хотели исследовать в естественных условиях, может ли лечение АЦ у мышей MDX также защищать от стресс-индуцированного повреждения. Первую серию экспериментов проводили при комнатной температуре. EDL мышцы изолированные от мышей всех групп прошли серию из четырех сокращений с измерением мышечной силы через 60 мин. В это время средняя сила для контрольных MDX мышц составила 53 ± 2% от первоначального значения (п = 6), в то время как мышцы от мышей, получавших лечение АЦ, имели значительно большую силу 60 ± 1% (п = 4, р <0,05 ). Во второй серии экспериментов при 35 ° С, силы достоверно не различалась между контролем и леченными MDX мышами (данные не показаны). Эти результаты показывают, что концентрация АЦ в мышцах мышей была достаточной для защиты от индуцированных АФК при комнатной температуре, но не при 35 ° С, где АФК намного больше (см. обсуждение).
АЦ предотвращает чрезмерную продукцию АФК в мышцах мышей MDX.
Для того, чтобы проверить, является ли лечение АЦ фактором снижения уровня АФК в мышцах MDX, мы использовали АФК-чувствительный краситель - ДГЕ. EDL сечения инкубировали с 5 мкм ДГЕ при 37 ° С, и измеряли интенсивность флуоресценции выше указанного порога. Как показано на рис. 4А, MDX мышцы контроля имели гораздо более широкое распространение и интенсивное окрашивание ядер, чем любые леченные MDX или контроль дикого типа мышц. Также может быть видно, что самое яркое окрашивание в контрольных мышцах MDX происходит в плотно упакованных областях, которые будут соответствовать проникновению воспалительных клеток, так как эти клетки могут производить значительное количество АФК (Nguyen & Tidball, 2003). Как показано на рис. 4B, площадь окрашивания в MDX мышцах 24,3 ± 4,4% (N = 9), которая значительно меньше (Р <0,05) в леченных MDX мышцах (7,5 ± 4,4%, п = 6, P <0,05) и контроле дикого типа (3,8 ± 1,4%, п = 5, р <0,01).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
