

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ПРАВИТЕЛЬСТВО РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ
УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗОВАНИЯ
«САНКТ-ПЕТЕРБУРГСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
(СПбГУ)
Кафедра физики атмосферы
Направление «Физика»

Гигроскопические и конденсационный свойства биологических аэрозолей
Магистерская диссертация студента
___________________
Научный руководитель:
__________________ д. ф.-м. н., проф.
Рецензент:
___________________ д. ф.-м. н., проф.
Санкт-Петербург
2018
Оглавление
Введение 3
Цель работы 3
Актуальность 3
Теория 6
Теория Келера 6
Параметр гигроскопичности 7
Дифференциальный анализатор подвижности 8
Техника эксперимента 11
Тандем дифференциальных анализаторов подвижности 11
Ошибки измерений 14
Метод подготовки и анализа клеточного материала 17
Результаты 19
Выводы 26
Список литературы 27
Введение
Цель работы
Биологический аэрозоль, то есть частицы растительного происхождения диаметром от нескольких нанометров до десятков микрон (например, пыльца растений, споры грибов, остатки растительности, бактерии и т. д.) составляет 15~25% глобальной аэрозольной массы. Как и любой другой аэрозоль, биочастицы участвуют во многих атмосферных процессах – рассеивают и поглощают свет, служат ядрами конденсации, а также являются сильными аллергенами. Но при этом, степень влияния биоаэрозолей на атмосферные процессы остается плохо изученной.
Целью научной работы является получение экспериментальных данных о механизмах взаимодействия биочастиц с атмосферным водяным паром, их гигроскопических свойств. Подобные данные можно использовать для упрощения прогнозирования результата их влияния на радиационное возмущение атмосферы.
Актуальность
Многочисленные лабораторные эксперименты показали, что первичные биочастицы являются активными ядрами конденсации и льдообразования и таким образом оказывают влияние на формирование облачных систем и осадков (Mцhler et al., 2007; Pцschl et al., 2010; DeMott et al., 2011; Morris et al., 2011; Hoose and Mцhler, 2012). Сравнительно недавно первичные биочастицы такие как споры грибов, бактерии были включены в климатические модели (Hoose et al., 2010; Spracklen et al., 2010; Sesartic et al., 2013). В целом авторы указывают, на то, что глобальный вклад первичных биочастиц (споры грибов, бактерии, пыльца растений) в формирование облачных систем незначителен (~1%). Тем не менее, авторы признают грубость модельных оценок, связанных, прежде всего, с ограниченностью данных об эмиссии и стоках биоаэрозолей и упрощенных подходах при учете их конденсационных и льдообразующих свойств. В частности, в модельных расчетах не учитывается вторичные субмикронные биочастицы, которые в большом количестве образуются в результате фрагментации исходных частиц. Так при резком изменении влажности пыльцевые зерна выделяют через поровые апертуры до ~104 субмикронных единиц клеточного материала (Solomon W. R., 2002; Grote et al., 2001; Tayler et al., 2002;2004; Grote et al., 2001) причем в среднем ~102-103 частиц являются водорастворимыми, способными к гигроскопическому росту (прямой форсинг) и служить облачными и ледяными ядрами конденсации (косвенный форсинг) (Pummer et al., 2015).
В целом, анализ литературных данных показывает, что в последнее десятилетие основные усилия исследователей были направлены на изучение льдообразующих свойств первичных биоаэрозолей (Blohn, 2005; Hoose et al., 2010; DeMott et al., 2011; Hoose and Mцhler, 2012; Augustin et al, 2013; Pummer et al., 2013; Adler et al., 2014; Frцhlich-Nowoisky et al., 2015). В меньшей степени изучены их гигроскопические и конденсационные свойства. В основном изучались конденсационные свойства бактерий и спор некоторых грибов (Pцschl et al.,2010; Pope, 2010; Mцhler et al., 2007). Что касается вторичных биоаэрозолей, то исследования их конденсационных свойств только начинают развиваться. На данный момент нам известна только одна работа (Steiner et al., 2015), в которой изучались конденсационные свойства вторичных продуктов пыльцевых зерен. Данные этой работы, в частности показывают, что конденсационная активность субчастиц (50-200 нм) цитоплазматического материала пыльцы (5 видов) сопоставимо с конденсационными свойствами сульфата аммония. Практически отсутствуют работы по гигроскопическим свойствам вторичных биологических аэрозолей.
Теория
Теория Келера
Равновесное давление пара ![]()
![]() над сферической каплей раствора при заданной температуре
над сферической каплей раствора при заданной температуре ![]()
![]() определяется из соотношения:
определяется из соотношения:
| (1) |
где ![]()
![]() – активность воды в капле раствора,
– активность воды в капле раствора, ![]()
![]() – молекулярная масса,
– молекулярная масса, ![]()
![]() – коэффициент поверхностного натяжения,
– коэффициент поверхностного натяжения, ![]()
![]() – плотность воды, R – универсальная газовая постоянная и
– плотность воды, R – универсальная газовая постоянная и ![]()
![]() – массовый (объемный) диаметр капли. Отношение давления пара над каплей к давлению насыщенного пара при заданной температуре
– массовый (объемный) диаметр капли. Отношение давления пара над каплей к давлению насыщенного пара при заданной температуре ![]()
![]() – степень насыщения. Если капля находится в равновесии с окружающим ее водяным паром, то эта величина может быть приравнена относительной влажности среды
– степень насыщения. Если капля находится в равновесии с окружающим ее водяным паром, то эта величина может быть приравнена относительной влажности среды ![]()
![]() , где
, где ![]()
![]() выражена в процентах. Уравнение (1) представляет собой одну из форм записи теории Келера (Koehler, 1936). Первое слагаемое
выражена в процентах. Уравнение (1) представляет собой одну из форм записи теории Келера (Koehler, 1936). Первое слагаемое ![]()
![]() описывает понижение давления водяного пара над плоской поверхностью в результате растворения вещества (поправка Рауля). Второе слагаемое
описывает понижение давления водяного пара над плоской поверхностью в результате растворения вещества (поправка Рауля). Второе слагаемое ![]()
![]() учитывает повышение давления пара над сферической каплей (поправка Кельвина,
учитывает повышение давления пара над сферической каплей (поправка Кельвина, ![]()
![]() ). В соотношении (1) слагаемое, учитывающее кривизну поверхности капли записано с учетом двух допущений. Предполагается, что поверхностное натяжение и парциальный мольный объем воды в растворе капли соответственно равны поверхностному натяжению
). В соотношении (1) слагаемое, учитывающее кривизну поверхности капли записано с учетом двух допущений. Предполагается, что поверхностное натяжение и парциальный мольный объем воды в растворе капли соответственно равны поверхностному натяжению ![]()
![]() и мольному объему
и мольному объему ![]()
![]() чистой воды. Данные расчетов показывают, что для частиц размером больше 100 нм эти упрощающие предположения приводят к ошибке определения равновесной относительной влажности меньше, чем 0.1% (Mikhailov et al., 2004).
чистой воды. Данные расчетов показывают, что для частиц размером больше 100 нм эти упрощающие предположения приводят к ошибке определения равновесной относительной влажности меньше, чем 0.1% (Mikhailov et al., 2004).
Для количественного описания гигроскопического роста частиц часто используется безразмерная величина, называемая фактором роста частиц, ![]()
![]() , которая определяется как
, которая определяется как
| (2) |
где ![]()
![]() – объемный диаметр сухой частицы. Соотношения (1) и (2) позволяют описать конденсационное укрупнение частиц при известных значениях активности воды в капле
– объемный диаметр сухой частицы. Соотношения (1) и (2) позволяют описать конденсационное укрупнение частиц при известных значениях активности воды в капле ![]()
![]() .
.
В работе (Kreidenweis et al., 2005) предложено физически обоснованное соотношение для описания зависимости фактора роста частиц от активности воды:
| (3) |
где ![]()
![]() ,
, ![]()
![]() и
и ![]()
![]() – параметры сглаживания. В данной работе эта форма использована для параметризации экспериментальных данных. Эффект влияния диаметра частиц на конденсационное равновесие был исключен подстановкой экспериментальных зависимостей
– параметры сглаживания. В данной работе эта форма использована для параметризации экспериментальных данных. Эффект влияния диаметра частиц на конденсационное равновесие был исключен подстановкой экспериментальных зависимостей ![]()
![]() в уравнение Келера (1) (Михайлов и др., 2008).
в уравнение Келера (1) (Михайлов и др., 2008).
Параметр гигроскопичности
Гигроскопические свойства аэрозольных частиц могут быть приблизительно описаны одним параметром гигроскопичности ![]()
![]() (Petters and Kreidenweis, 2007):
(Petters and Kreidenweis, 2007):
| (4) |
С учетом аддитивности объемов, уравнение (4) можно переписать как:
| (5) |
Дифференциальный анализатор подвижности
Электрическая подвижность является мерой способности частицы двигаться в электрическом поле. Электрическая подвижность ![]()
![]() определяется уравнением:
определяется уравнением:
| (6) |
где n – число элементарных зарядов на частице, e – элементарный заряд (![]()
![]() ), C – поправка скольжения Каннингема,
), C – поправка скольжения Каннингема, ![]()
![]() – вязкость газа,
– вязкость газа, ![]()
![]() – диаметр частицы.
– диаметр частицы.
Принцип работы дифференциального анализатора подвижности (Differential Mobility Analyzer, DMA) основан на электрической подвижности. DMA представляет собой полый цилиндр со стержнем внутри, на них подается высокое напряжение (рис.1). Вдоль стержня идет постоянный поток воздуха, сверху вниз. Частицы аэрозоля попадают сверху, и идут вдоль цилиндра. То, как сильно будет отклоняться траектория частиц будет определятся электрической подвижностью. Выходная щель в нижней части DMA устроена таким образом, чтобы на выход попадала лишь часть частиц, отклонившихся на заданный угол, все остальные попадут либо на стенки, либо на стержень DMA и выйдут с избытком воздуха. Таким образом, при постоянной скорости потока частиц и напряжении DMA, на выход будут попадать только частицы с узким диапазоном электрической подвижности, который можно варьировать, меняя напряжение.

Рис. 1. Устройство дифференциального анализатора подвижности
Но этот диапазон диаметров частиц зависит не только от электрической подвижности частиц. Кнутсон (Knutson, 1975) определил взаимосвязь между электрической подвижностью частиц ![]()
![]() и параметрами прибора и ее ошибку
и параметрами прибора и ее ошибку ![]()
![]() :
:
| (7) |
| (8) |
где ![]()
![]() – расход аэрозоля,
– расход аэрозоля, ![]()
![]() – расход воздуха, идущего через DMA,
– расход воздуха, идущего через DMA, ![]()
![]() и
и ![]()
![]() – внешний и внутренний радиус кольцевой полости,
– внешний и внутренний радиус кольцевой полости, ![]()
![]() – среднее напряжение на стержне,
– среднее напряжение на стержне, ![]()
![]() – расстояние от входа полидисперсного аэрозоля до выходной щели. Объединив выражения (6) и (7), можно получить уравнение, которое будет связывать диаметр частиц с напряжением на стержне, количеством зарядов на частице, расходом потока воздуха и геометрией самого прибора (Series 3080 Electrostatic Classifiers Operation and Service Manual, 2009):
– расстояние от входа полидисперсного аэрозоля до выходной щели. Объединив выражения (6) и (7), можно получить уравнение, которое будет связывать диаметр частиц с напряжением на стержне, количеством зарядов на частице, расходом потока воздуха и геометрией самого прибора (Series 3080 Electrostatic Classifiers Operation and Service Manual, 2009):
| (9) |
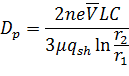
Техника эксперимента
Тандем дифференциальных анализаторов подвижности
Работу тандема дифференциальных анализаторов подвижности (ТДАП, Hygroscopicity Tandem Differential Mobility Analyzer, HTDMA) можно условно поделить на три части: генерация и выделение частиц, их взаимодействие с водяным паром, определение диаметра. Полная схема показана на рис. 2.
Рис. 2. Схема ТДАП
I. Сначала раствор воды с частицами аэрозоля распыляется форсункой. Вещества приготавливались в виде раствора в деионизованной воде с удельным сопротивлением 18.2 МОмсм. Мелкие капельки проходят через диффузионный осушитель, наполненный силикагелем, который поглощает всю воду. Получившийся полидисперсный аэрозоль проходит нейтрализатор заряда и попадает в первый дифференциальный анализатор подвижности (Differential Mobility Analyzer, DMA). Первый DMA выделяет из сухого полидисперсного аэрозоля с очень узким диапазоном диаметров. На рис. 3 показан пример того, как выглядит результат этого отбора.
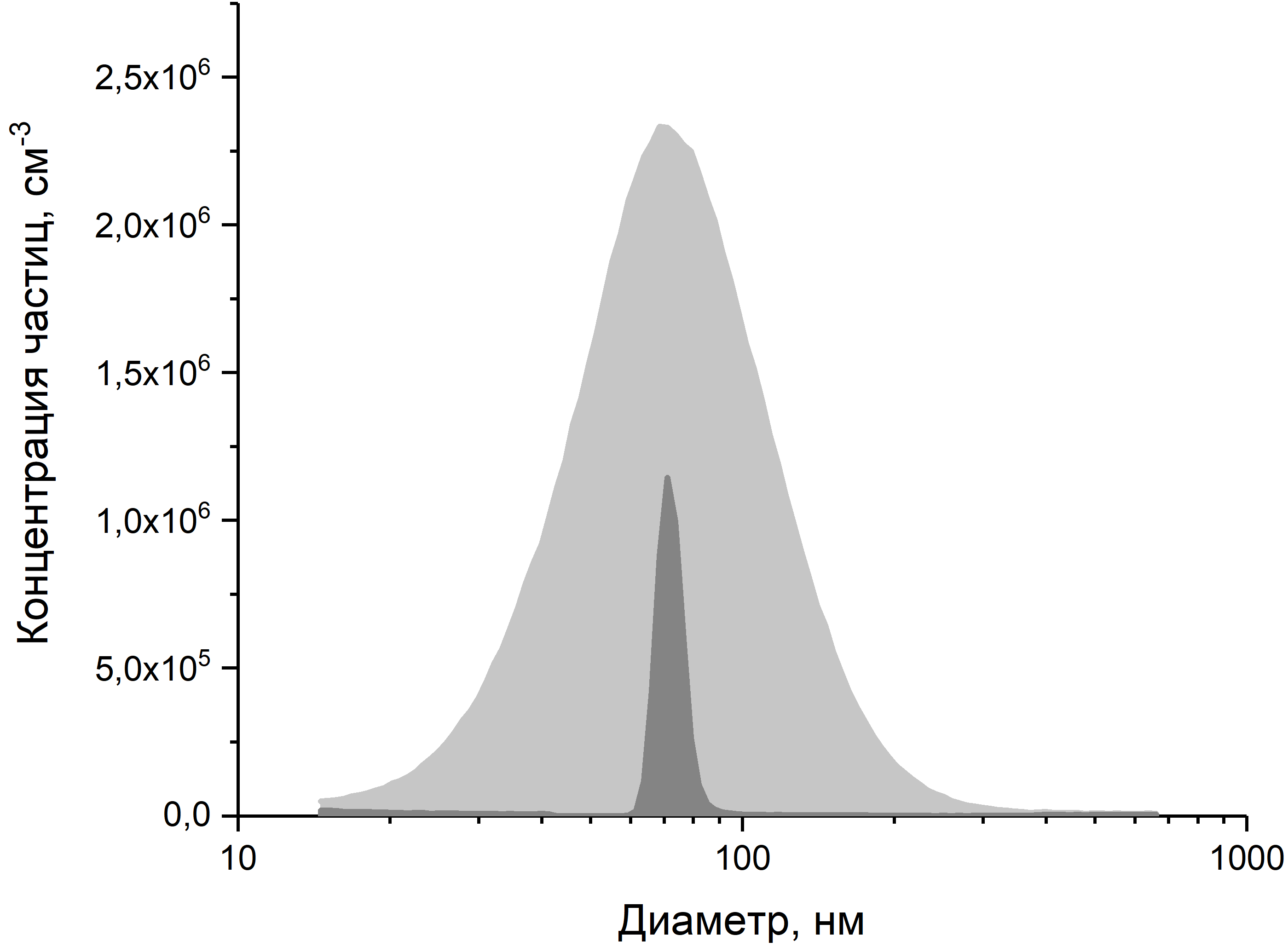
Рис. 3. Пример того, как первый DMA выделяет узкий монодисперсный диапазон частиц (темная область) из общего полидисперсного аэрозоля (светлая область).
II. Взаимодействии аэрозоля с водяным паром проходило в трех режимах.
A. Режим гидратации, возрастающей влажности. Сухие частицы попадают в увлажнитель. Он представляет собой двухслойную трубку, внешний слой непроницаемый, а внутренний способен пропускать молекулы воды, поскольку выполнен из полупроницаемого материала Nafion. Между слоями идет воздух при заданной относительной влажности (она достигается смешиванием сухого и влажного воздуха в разных пропорциях). Таким образом, когда аэрозоль идет по внутренней трубке, он смешивается с парами воды при заданной относительной влажности. Время увлажнения аэрозоля составляет около 13 секунд, что исключает влияние кинетического фактора на конденсационный рост (Седунов, 1972; Kerminen, 1997).
B. Режим дегидратации, убывающей влажности. Его отличие от гидратации в том, что перед увлажнителем находится еще один, с таким же устройством, но у него пространство между слоями полностью заполнено водой. То есть после него аэрозоль имеет около 94% относительной влажности. Далее он так же, попадает в увлажнитель.
C. Режим реструктуринга. Так же, как и в режиме гидратации аэрозоль проходит увлажнитель, но после этого он также проходит осушитель, в котором силикагель поглощает всю влагу. Этот режим используется для того, чтобы выявить эффект реструктуринга.
III. Аэрозоль попадает на второй DMA, который в отличии от первого, является сканирующим анализатором подвижности, т. е. напряжение меняется во время эксперимента. После аэрозоль попадает на счетчик частиц, который измеряет концентрацию. В ходе каждого измерения, в течении некоторого времени аэрозоль проходит второй DMA, который сканирует по заданному диапазону диаметров, например, от 70 до 150 нм. Диаметры сопоставляются с концентрацией частиц в каждый момент времени, что и позволяет понять, каким образом происходит гигроскопический рост аэрозоля при взаимодействии с водяным паром.
На практике для расчета фактор роста использовано соотношение:
| (10) |
где ![]()
![]() и
и ![]()
![]() – эквивалентный диаметр сухой частицы по объему и по подвижности, соответственно, D – влажный диаметр частицы и
– эквивалентный диаметр сухой частицы по объему и по подвижности, соответственно, D – влажный диаметр частицы и ![]()
![]() – фактор реструктуринга, параметр учитывающий отличие формы исходной частицы от сферической. Его связь с динамическим фактором формы частицы
– фактор реструктуринга, параметр учитывающий отличие формы исходной частицы от сферической. Его связь с динамическим фактором формы частицы ![]()
![]() определяется из соотношения:
определяется из соотношения:
| (11) |
где ![]()
![]() – поправка Канингема, учитывающая увеличение подвижности частиц для помежуточных чисел Кнудсона (~1-10). Парметр реструктуринга
– поправка Канингема, учитывающая увеличение подвижности частиц для помежуточных чисел Кнудсона (~1-10). Парметр реструктуринга ![]()
![]() определяется по данным ТДАП измерений в режиме реструктурига (С), предполагая, что
определяется по данным ТДАП измерений в режиме реструктурига (С), предполагая, что ![]()
![]() , т. е.
, т. е.![]()
![]() .
.
Ошибки измерений
- Активность воды
- Гигроскопический параметр
- Относительная влажность
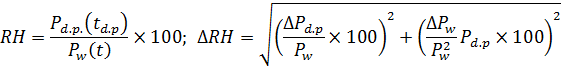
- Фактор роста
Ошибка определения фактора роста зависит от параметров дифференциального анализатора подвижности, а именно – от неопределенности расхода воздуха, изменении геометрии DMA анализатора вследствие осаждения частиц на его внутренних электродах и вариации напряжения (см. формулу 9). В нашем эксперименте относительная ошибка измерений фактора роста ![]()
![]() составила 0.02.
составила 0.02.
На рис. 4 и 5 показаны калибровочные данные измерений сульфата аммония. На рис. 4 видно, что экспериментальные данные хорошо сходятся с данными, рассчитанными по модели Келера для сульфата аммония. На рис. 5 показано, что даже при высокой относительной влажности, ошибки в допустимом пределе.
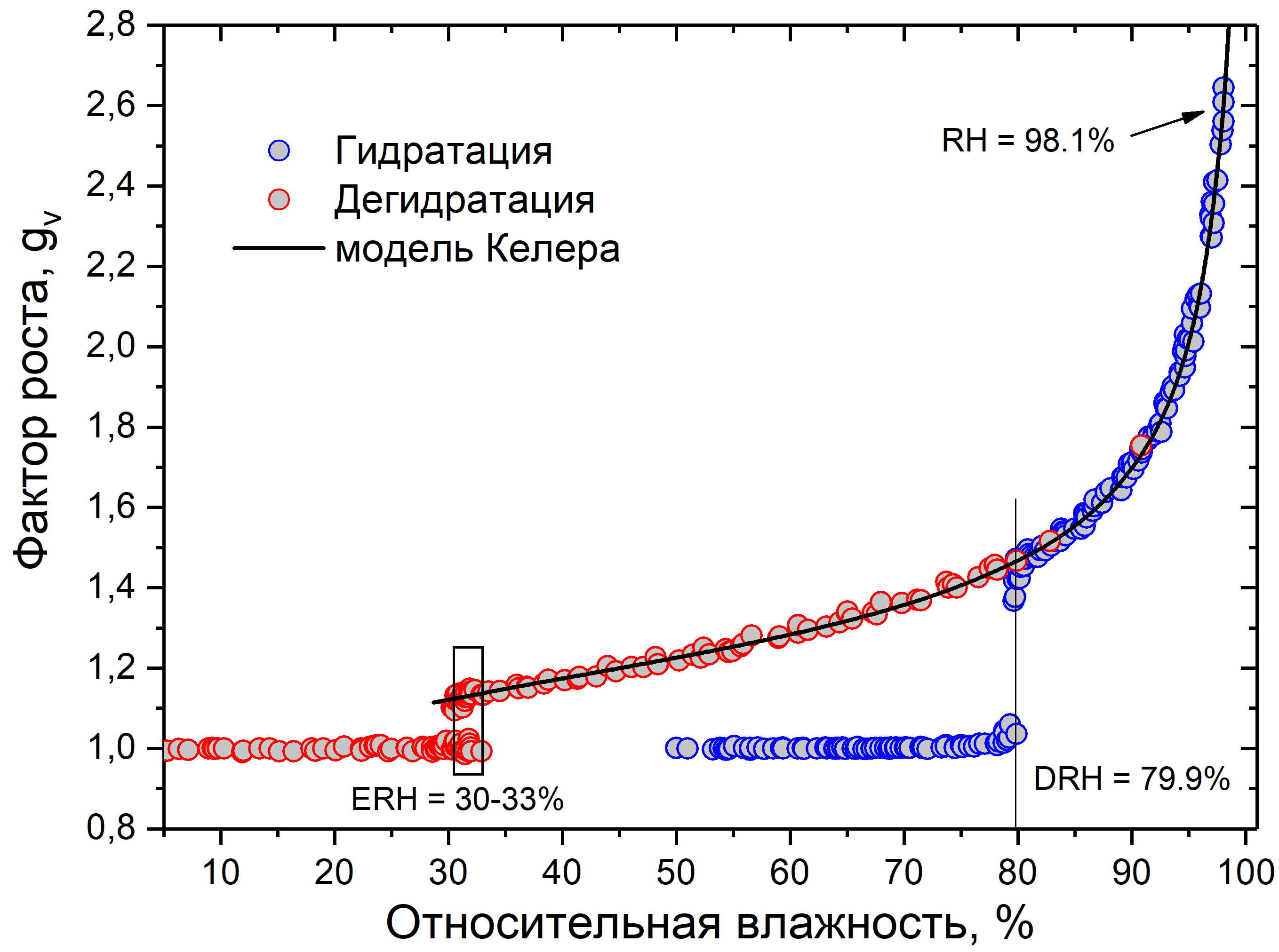
Рис. 4. График зависимости фактора роста от относительной влажности для сульфата аммония
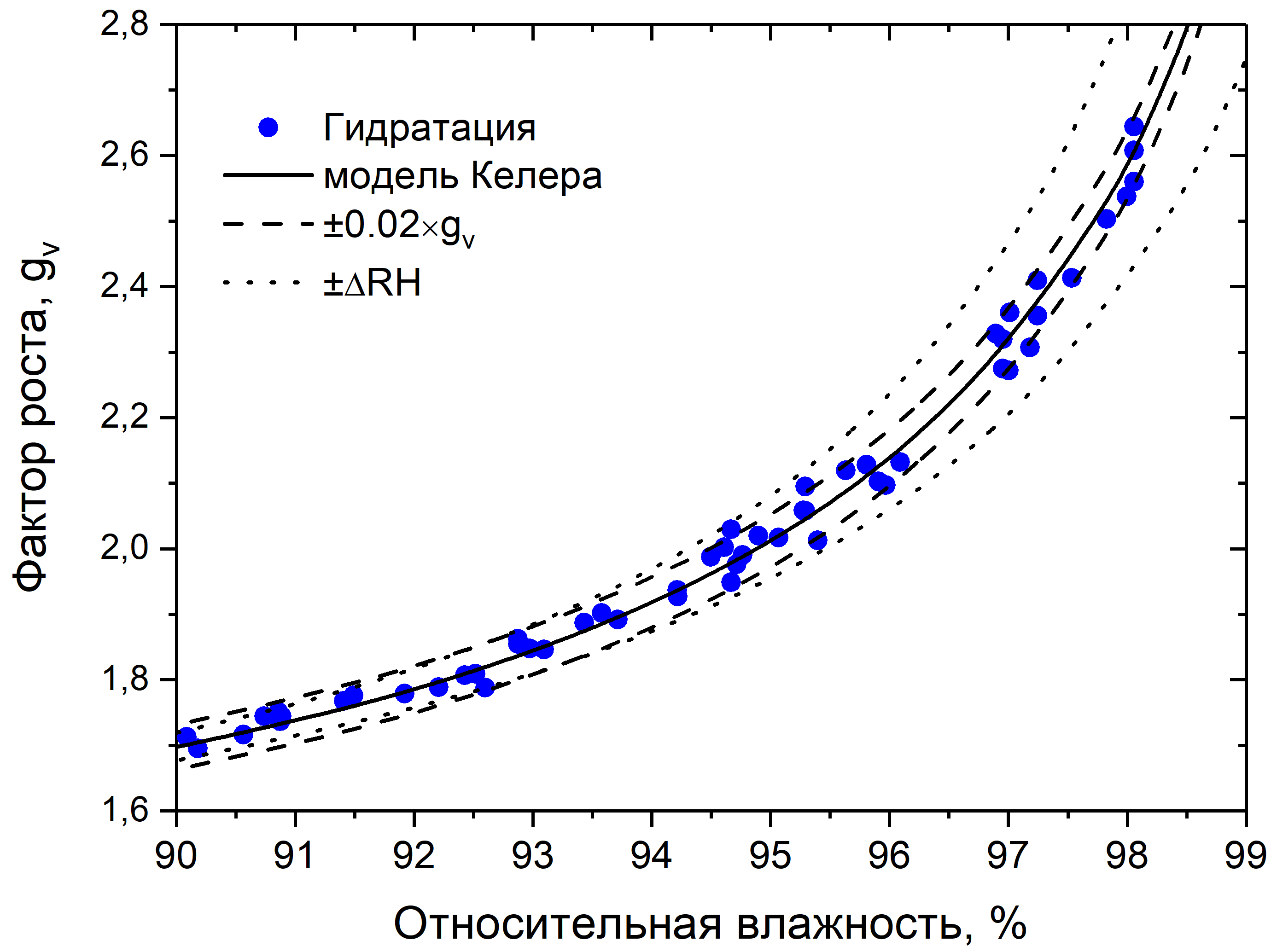
Рис. 5. График зависимости фактора роста от относительной влажности (в диапазоне 90-99%) для сульфата аммония, с погрешностями
Метод подготовки и анализа клеточного материала
Проводилось исследование органического состава водорастворимой фракции биоаэрозолей. Разработана методика анализа основных компонентов клеточного материала пыльцевых зерен. Эта методика включает:
Экстракцию водорастворимых компонентов из пыльцевых гранул с помощью ультразвуковой ванны в течение 40 минут (рис. 6).
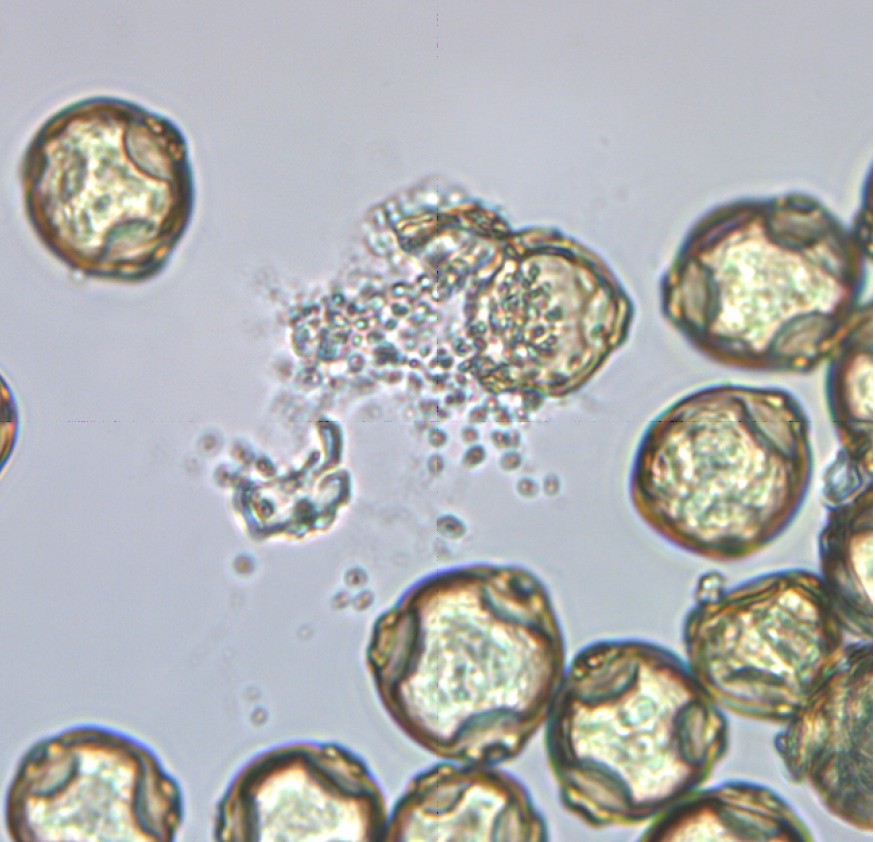
Рис. 6. Пыльцевые гранулы, выпускающие клеточный материал
Фильтрацию раствора, т. е. его очистку от нерастворимых в воде материалов с помощью нуклеопорных фильтров (8 мкм и 0.45 мкм).
Был проведен анализ водных растворов на содержание в них протеина, углеводов и нуклеиновых кислот (рис. 7).
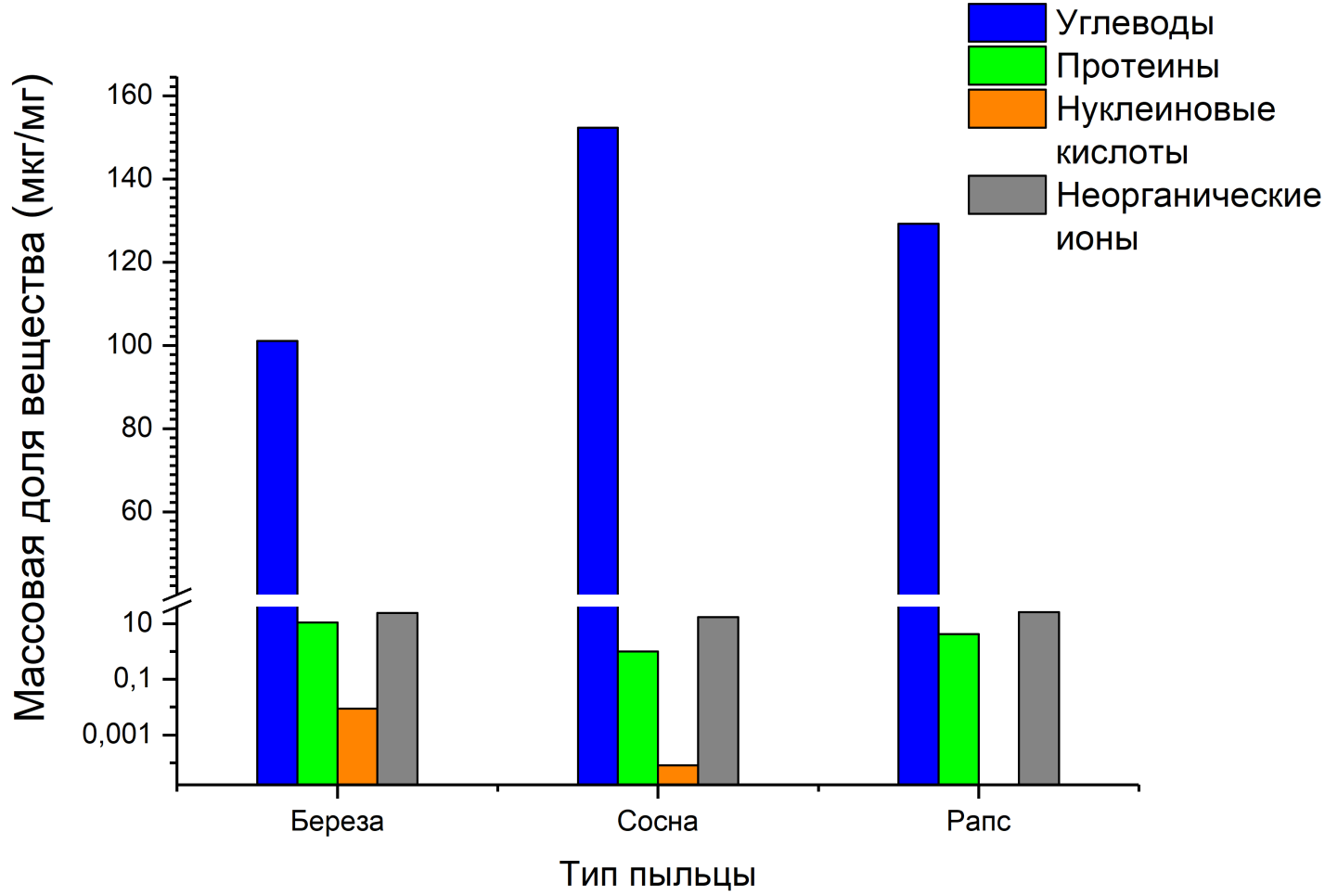
Рис. 7. Результаты анализа водных растворов на содержание в них протеина, углеводов, нуклеиновых кислот и неорганических ионов
Содержание растворимых углеводов в пробах определялось фенол-сульфатным методом, описанным в работе (Masuko et. al. 2005). Для определения содержания протеинов в пробах использовано 2 метода: стандартный «Qubit protein assay» (Thermo fisher Scientific) и хим. анализ аминокислот. Оба метода дали близкие результаты (в пределах 10%). Общее содержание нуклеиновых кислот в пробах проводилось также по стандартной методике «Qubit DNA assay» (Thermo fisher Scientific). Метод ионной хроматографии использован для определения содержания в пробах неорганических ионов F, Cl, NO2, Br, NO3, PO4, SO4, Na, NH4, K, Ca и Mg.
Представленные на рис. 5 данные аналитических измерений указывают на то, что основными водопоглощающими компонентами пыльцевых зерен являются водорастворимые углеводы, протеины и неорганические ионы.
Результаты
На рис. 8-10 представлены данные измерений для сосны, березы и рапса. Каждый график показывает зависимость фактора роста от активности воды в режимах гидратации и дегидратации, для частиц 70 и 100 нм. Также на каждый график нанесено сглаживание с использованием модели Келера, построенное по формуле (5). Коэффициенты сглаживания в табл. 1.
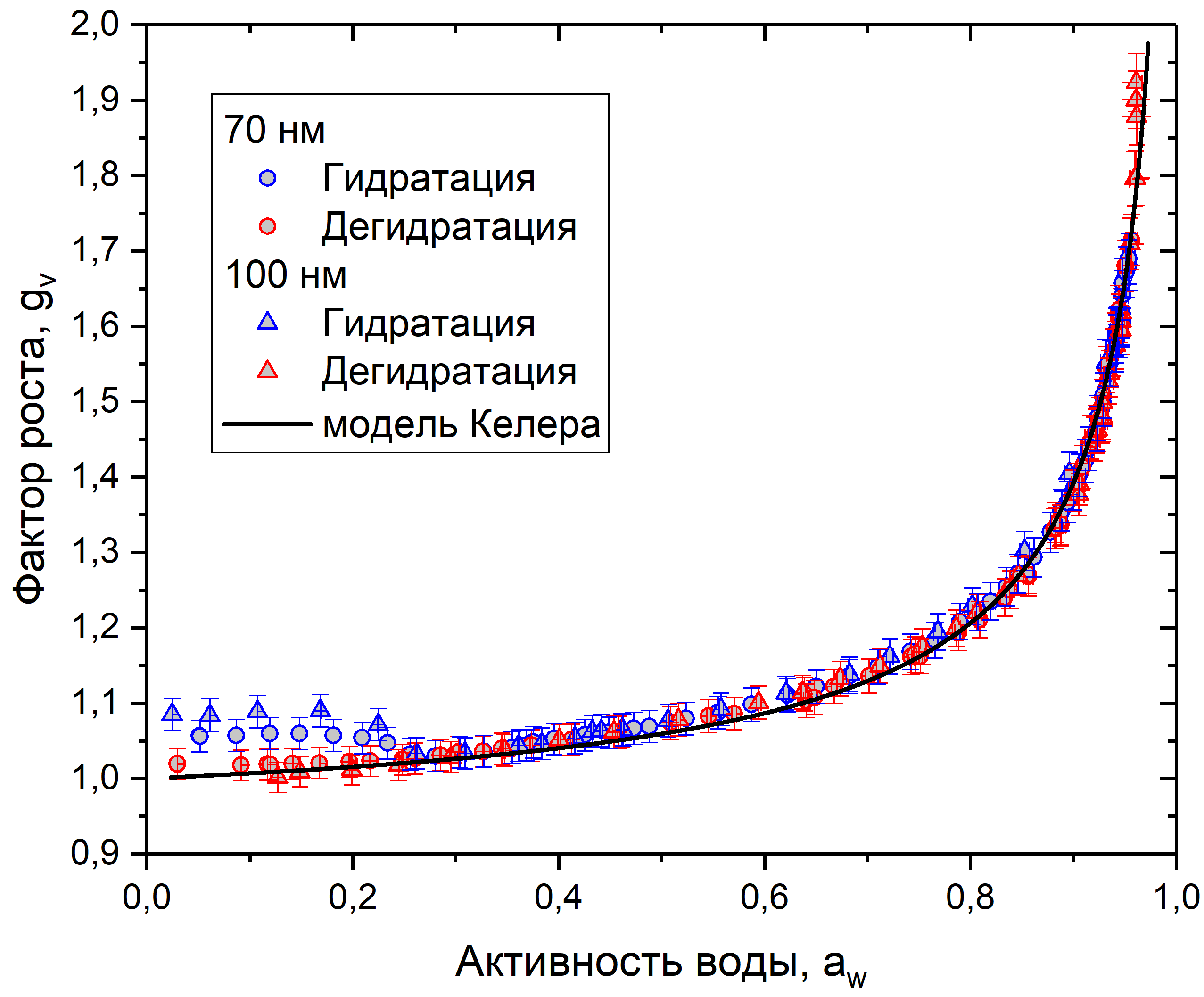
Рис. 8. График зависимости фактора роста от активности воды для клеточного материала пыльцы сосны
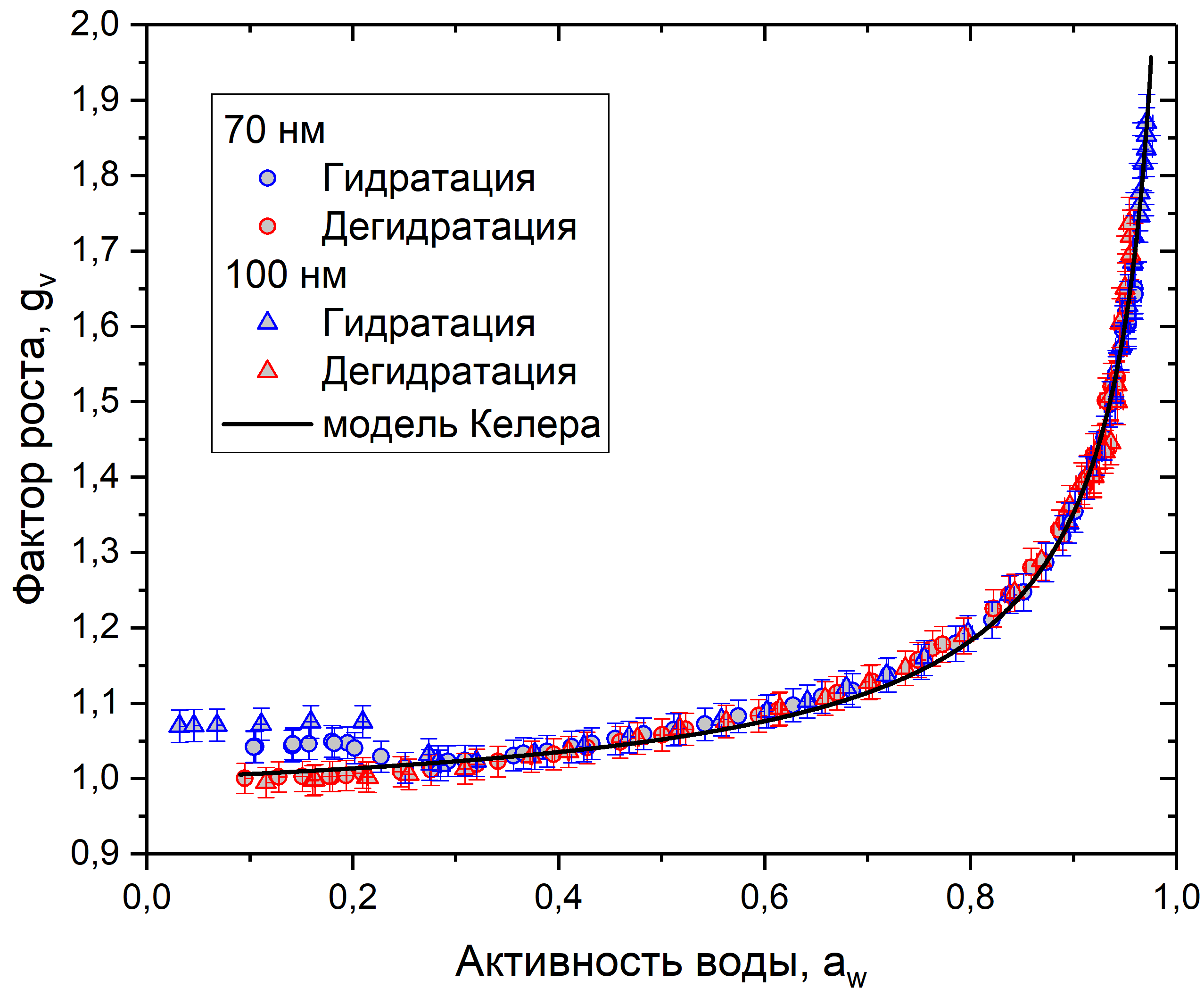
Рис. 9. График зависимости фактора роста от активности воды для клеточного материала пыльцы березы
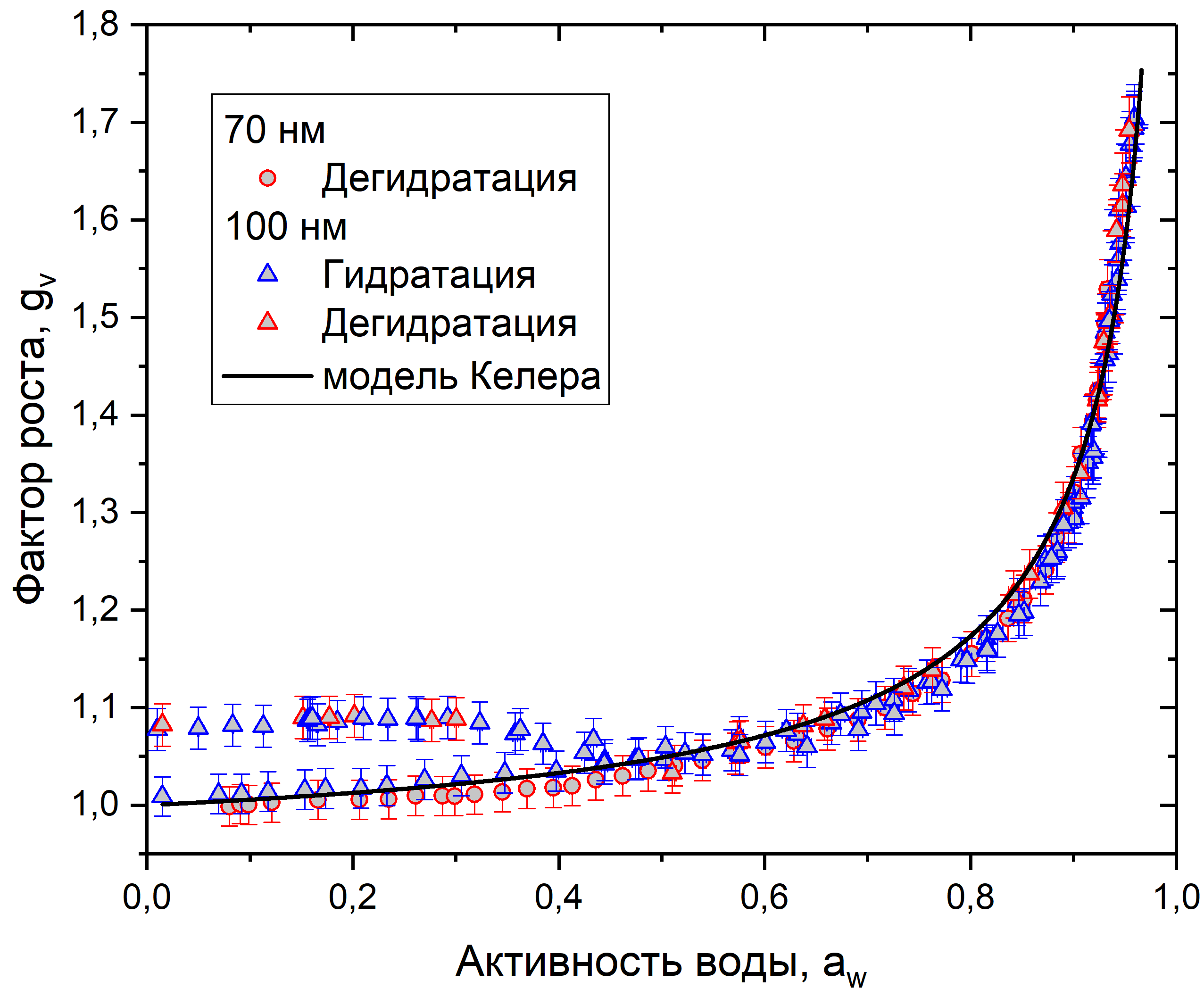
Рис. 10. График зависимости фактора роста от активности воды для клеточного материала пыльцы рапса
На рис. 11-13 представлены данные измерений для сосны, березы и рапса. Каждый график показывает зависимость гигроскопического фактора от активности воды. Также на каждый график нанесено сглаживание с использованием формулы ![]()
![]() . Коэффициенты сглаживания представлены в табл. 2.
. Коэффициенты сглаживания представлены в табл. 2.
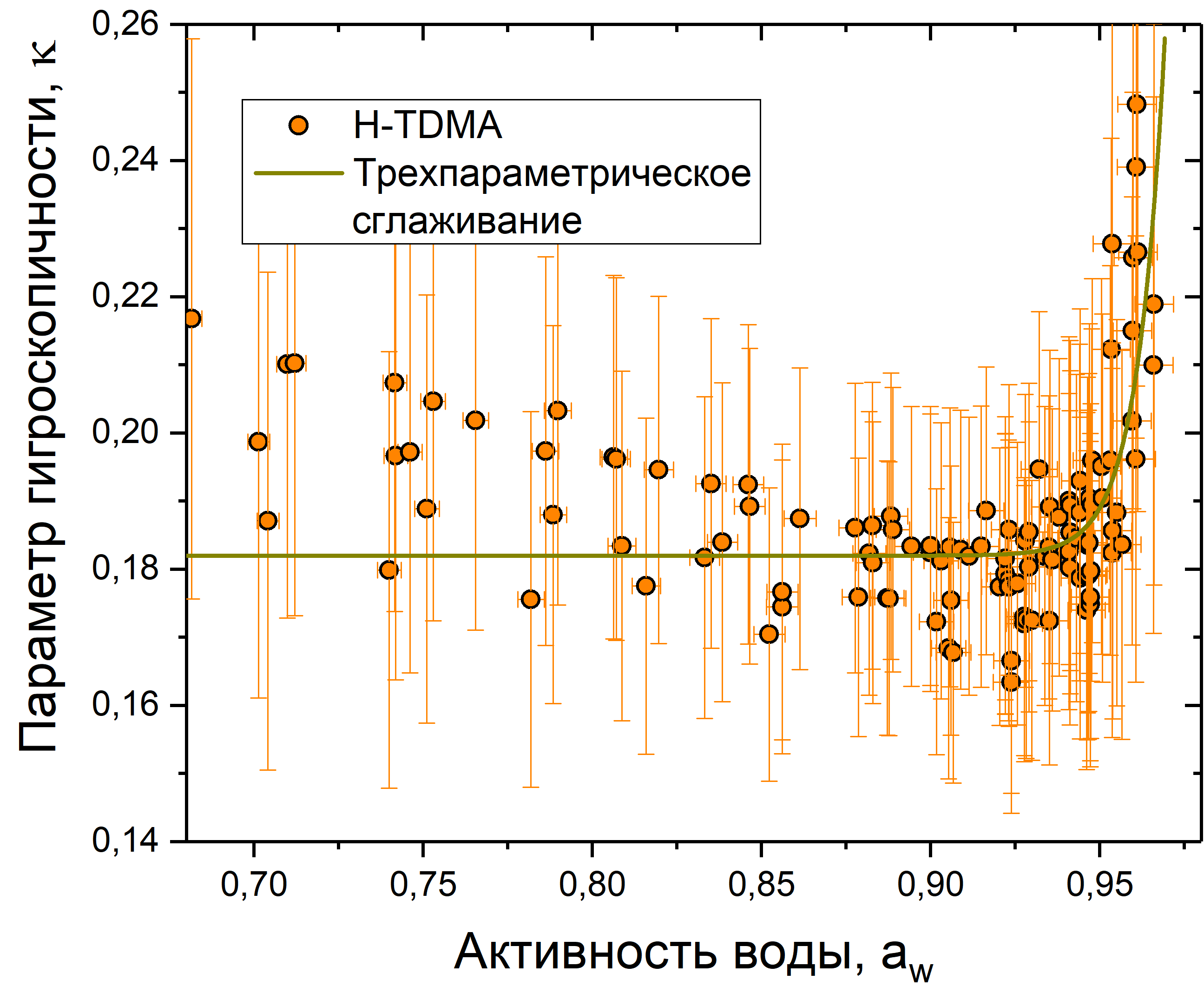
Рис. 11. График зависимости параметра гигроскопичности от активности воды для клеточного материала пыльцы сосны

Рис. 12. График зависимости параметра гигроскопичности от активности воды для клеточного материала пыльцы березы
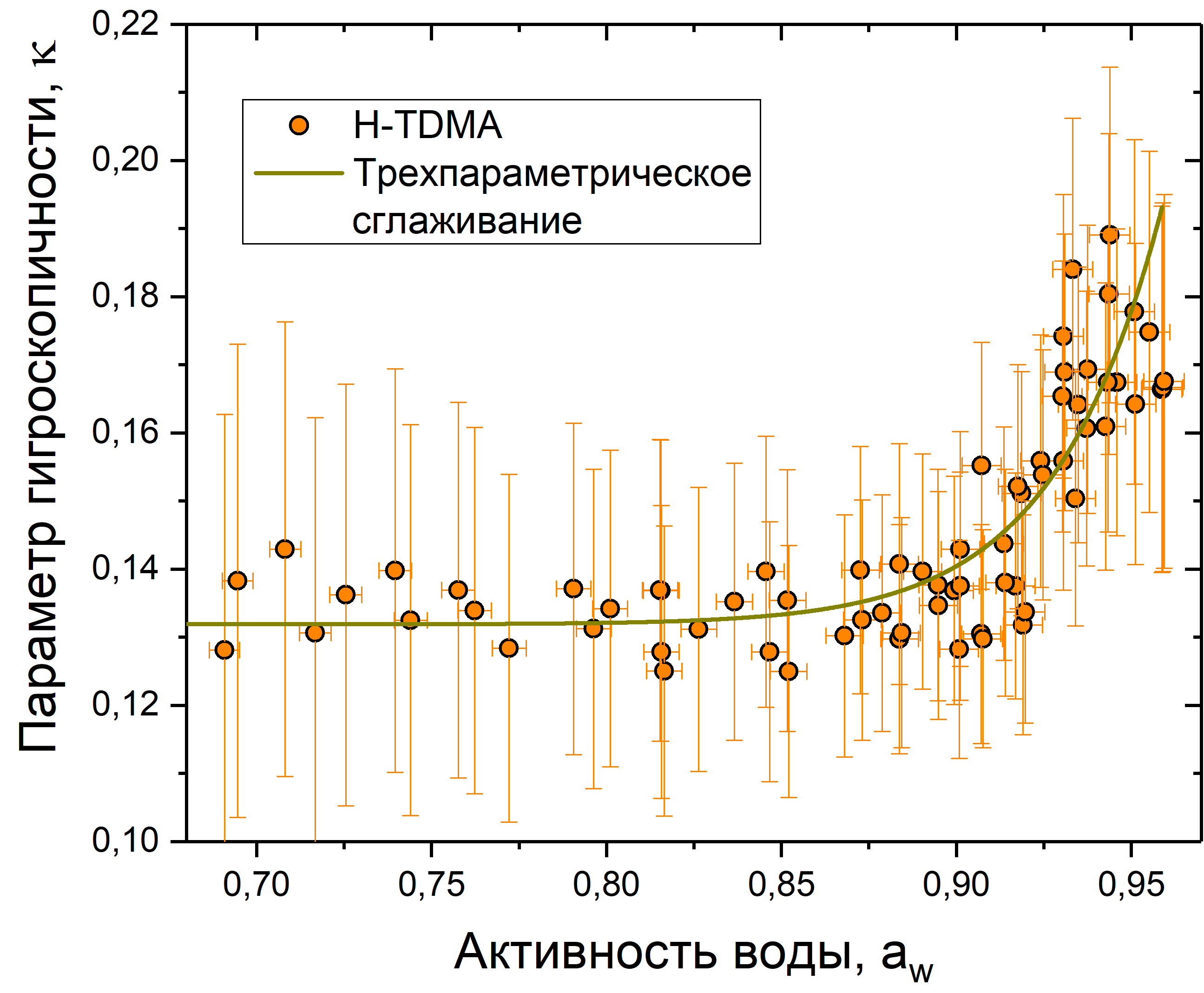
Рис. 13. График зависимости параметра гигроскопичности от активности воды для клеточного материала пыльцы рапса
| |
Сосна | 0.189 |
Береза | 0.164 |
Рапс | 0.154 |
Табл. 1. Коэффициенты сглаживания ![]()
![]()
a | b | c | |
Сосна | 0.182 | 3.127 | 118.913 |
Береза | 0.239 | -0.085 | 1.380 |
Рапс | 0.132 | 0.227 | 31.138 |
Табл. 2. Коэффициенты сглаживания ![]()
![]()
Выводы
Была разработана и усовершенствована методика генерации, подготовки и определения диаметра пыльцевых зерен в трех режимах – гидратации, дегидратации и реструктуринга. Работа тандема дифференциальных анализаторов подвижности была усовершенствована, температурные градиенты во всей системе были практически полностью устранены, введен более точный способ определения относительной влажности с использованием новых приборов.
Были получены уникальные данные по гигроскопическому росту и свойствам для трех типов пыльцы – сосны, березы и рапса. Подобные данные можно использовать для упрощения прогнозирования результата их влияния на радиационное возмущение атмосферы.
Список литературы
1. Mцhler, O., DeMott, P. J., Vali, G., and Levin, Z.: Microbiology and atmospheric processes: the role of biological particles in cloud physics, Biogeosciences, 4, 1059-1071, https://doi. org/10.5194/bg-4-1059-2007, 2007.
2. Pцschl, U., Martin, S. T., Sinha, B., et al.: Rainforest Aerosols as Biogenic Nuclei of Clouds and Precipitation in the Amazon, Science, 329, p.1513-1515, 2010. DOI: 10.1126/science.
3. DeMott, P. J., O. Mцhler, O. Stetzer, G. Vali, Z. Levin, M. D. Petters, M. Murakami, T. Leisner, U. Bundke, H. Klein, Z. A. Kanji, R. Cotton, H. Jones, S. Benz, M. Brinkmann, D. Rzesanke, H. Saathoff, M. Nicolet, A. Saito, B. Nillius, H. Bingemer, J. Abbatt, K. Ardon, E. Ganor, D. G. Georgakopoulos, and C. Saunders, 2011: Resurgence in Ice Nuclei Measurement Research. Bull. Amer. Meteor. Soc., 92, 1623–1635, https://doi. org/10.1175/2011BAMS3119.1 .
4. Morris C. E., Conen F., Huffman J. A.: Bioprecipitation: a feedback cycle linking Earth history,
5. ecosystem dynamics and land use through biological ice nucleators in the atmosphere, Global Change Biology 20, 341–351, doi: 10.1111/gcb.12447, 2014.
6. Hoose, C. and Mцhler, O.: Heterogeneous ice nucleation on atmospheric aerosols: a review of results from laboratory experiments, Atmos. Chem. Phys., 12, 9817-9854, https://doi. org/10.5194/acp-12-9817-2012, 2012.
7. Hoose C., Kristjansson J. E. and Burrows S. M.. How important is biological ice nucleation in clouds on a global scale? Environ. Res. Lett. 5: 024009, doi:10.1088/1748-9326/5/2/024009, 2010.
8. Spracklen, D. V., Carslaw, K. S., Merikanto, J., Mann, G. W., Reddington, C. L., Pickering, S., Ogren, J. A., Andrews, E., Baltensperger, U., Weingartner, E., Boy, M., Kulmala, M., Laakso, L., Lihavainen, H., Kivekдs, N., Komppula, M., Mihalopoulos, N., Kouvarakis, G., Jennings, S. G., O'Dowd, C., Birmili, W., Wiedensohler, A., Weller, R., Gras, J., Laj, P., Sellegri, K., Bonn, B., Krejci, R., Laaksonen, A., Hamed, A., Minikin, A., Harrison, R. M., Talbot, R., and Sun, J.: Explaining global surface aerosol number concentrations in terms of primary emissions and particle formation, Atmos. Chem. Phys., 10, 4775-4793, https://doi. org/10.5194/acp-10-4775-2010, 2010.
9. Sesartic A., Lohmann U. & Storelvmo T. 2013. Modelling the impact of fungal spore ice nuclei on clouds and precipitation. Environ. Res. Lett. 8: 014029, doi:10.1088/1748-9326/8/1/014029, 2013.
10. Solomon W. R.: Airborne pollen: A brief life, J Allergy Clin. Immunol., June, 895-900, 2002.
11. Grote M., Vrtala S., Niederberger V. et al.: Release of allergen-bearing cytoplasm from hydrated pollen: A mechanism common to a variety of grass (Poaceae) species revealed by electron microscopy, J. Allerg. Clin. Immunol., 108, 109–115, doi:10.1067/mai.2001.116431, 2001.
12. Taylor PE, Flagan RC, Valenta R, Glovsky MM. Release of allergens as respirable aerosols: A link between grass pollen and asthma. J Allergy Clin Immunol. 2002 Jan;109(1):51-6. PubMed PMID: 11799365.
13. Taylor PE, Flagan RC, Miguel AG, Valenta R, Glovsky MM. Birch pollen rupture and the release of aerosols of respirable allergens. Clin Exp Allergy. 2004 Oct;34(10):1591-6. PubMed PMID: 15479275.
14. Pummer, B. G., Budke, C., Augustin-Bauditz, S., Niedermeier, D., Felgitsch, L., Kampf, C. J., Huber, R. G., Liedl, K. R., Loerting, T., Moschen, T., Schauperl, M., Tollinger, M., Morris, C. E., Wex, H., Grothe, H., Pцschl, U., Koop, T., and Frцhlich-Nowoisky, J.: Ice nucleation by water-soluble macromolecules, Atmos. Chem. Phys., 15, 4077-4091, https://doi. org/10.5194/acp-15-4077-2015, 2015.
15. Blohn N., Mitra S. K., Diehl et al.: The ice nucleating ability of pollen Part III: New laboratory studies in immersion and contact freezing modes including more pollen types, Atmospheric Research 78, 182– 189, 2005.
16. Augustin, S., Wex, H., Niedermeier, D., Pummer, B., Grothe, H., Hartmann, S., Tomsche, L., Clauss, T., Voigtlдnder, J., Ignatius, K., and Stratmann, F.: Immersion freezing of birch pollen washing water, Atmos. Chem. Phys., 13, 10989-11003, https://doi. org/10.5194/acp-13-10989-2013, 2013.
17. Pummer, B. G., Atanasova, L., Bauer, H., Bernardi, J., Druzhinina, I. S., Frцhlich-Nowoisky, J., and Grothe, H.: Spores of many common airborne fungi reveal no ice nucleation activity in oil immersion freezing experiments, Biogeosciences, 10, 8083-8091, https://doi. org/10.5194/bg-10-8083-2013, 2013.
18. Adler G., Koop N., Haspel C. et al.: Formation of highly porous aerosol particles by atmospheric freeze-drying in ice clouds, PNAS, 110, 20414-20419, 2013.
19. Frцhlich-Nowoisky J., Hill T. C., Pummer B. G. et al.: Ice nucleation activity in the widespread soil fungus Mortierella alpine, Biogeosciences, 12, 1057–1071, 2015.
20. Pope F. D.: Pollen grains are efficient cloud condensation nuclei, Environ. Res. Lett. 5, 044015 (6pp), (2010).
21. Steiner, A. L., Brooks, S. D. Deng C., et al.: Pollen as atmospheric cloud condensation nuclei, Geophys. Res. Lett., 42, 3596–3602, doi:10.1002/2015GL064060, 2015.
22. Koehler H. The nucleus in and the growth of hydroscopic droplets // Trans. Farad. Soc. 1936. V. 32. P. 1152–1161, doi: 10.1039/TF9363201152.
23. Mikhailov E., Vlasenko S., Niessner R., Pцschl U. Interaction of aerosol particles composed of protein and salts with water vapor: hygroscopic growth and microstructural rearrangement, Atmos. Chem. Phys. 2004. V. 4. ‹ 2. P. 323–350.
24. Kreidenweis S. M., Koehler K., DeMott P. et al. Water activity and activation diameters from hygroscopic data. Pt I: theory and application to inorganic salts, Atmos. Chem. Phys. 2005. V. 5. ‹ 5. P. 1357–1370.
25. Михайлов, Е. Ф., Власенко, С. С., & Рышкевич, Т. И. (2008). ВЛИЯНИЕ ХИМИЧЕСКОГО СОСТАВА И МИКРОСТРУКТУРЫ НА ГИГРОСКОПИЧЕСКИЙ РОСТ ПИРОГЕННОГО АЭРОЗОЛЯ. ИЗВЕСТИЯ РОССИЙСКОЙ АКАДЕМИИ НАУК. ФИЗИКА АТМОСФЕРЫ И ОКЕАНА, 44(4), 450-466. DOI: DOI: 10.1134/S0002351508040032.
26. Petters, M. D. and Kreidenweis, S. M.: A single parameter representation of hygroscopic growth and cloud condensation nucleus activity, Atmos. Chem. Phys., 7, 1961-1971, https://doi. org/10.5194/acp-7-1961-2007, 2007.Knutson E. O., and K. T. Whitby Aerosol Classification by Electric Mobility: Apparatus Theory and Applications, Journal of Aerosol Science, 1975, 6:443.
27. Series 3080 Electrostatic Classifiers Operation and Service Manual, P/N 1933792, Revision J, March 2009.
28. Физика образования жидко-капельной фазы в атмосфере. Л.: Гидрометеоиздат, 1972. 207 с.
29. Kerminen V. M. The effects of particle chemical character and atmospheric processes on particle hygroscopic properties, J. Aeros. Sci. 1997. V. 28. P. 121–132.
30. Masuko T, Minami A, Iwasaki N, Majima T, Nishimura S, Lee YC. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal Biochem 2005 Apr 1;339(1):69-72. PubMed PMID: 15766712.
