
Окрашивание синим красителем Эванса и креатинкиназа сыворотки.
10 мг / мл раствор голубого Эванса красителя в PBS инъецировали в хвостовую вену мышей дикого типа, бигликан - дефицитных и MDX мышей. Четырехглавые мышцы были собраны через 3-6 ч и сделаны криосекции. Синий краситель Эванса - положительные мышечные волокна наблюдались под флуоресцентным микроскопом, используя набор родаминовых фильтров. Сто мышечных волокон в секции для каждой из трех мышей на генотип были подсчитаны. Для измерений КФК в сыворотке собирали кровь анестезированных животных ретро-орбитального кровотечения со сгустками, а уровни фермента были измерены с использованием коммерческого набора (Sigma) .
Гистология и иммуногистохимия
Четырехглавая мышца бедра была выделена и замораживали в жидком изопентане охлажденном азотом. Во всех экспериментах 10 мкм срезы ровесников конгенных BGN ноль и дикого типа мышей были установлены на том же слайде. Срезы фиксировали 4% параформальдегидом свежеприготовленным в течение 5 мин при комнатной температуре, блокированной в блокирующем реагенте в течение 1 ч при комнатной температуре и инкубировали с указанными первичными Aт ночь при 4 ° С с последующей инкубацией с вторичными Aт в течение 1 ч при комнатной температуре.
Вектор M. O.M.
Базовый комплект использовали в соответствии с протоколом производителя. Срезы установлены в Permafluor ( Thermo Electron Corporation, Pittsburgh, PA, США) и анализировали с помощью конфокальной лазерной сканирующей микроскопии ( Leica TCS SP2 акустооптического светоделитель ( AOBS ) . Иммунного окрашивание не наблюдалось, IgG или значения Aт были заменены первым слоем; исследование также показало, что вторичные антитела видоспецифичны. Все изображения для сравнения, представленные здесь, были захвачены со срезов BGN нуль и дикого типа мышц, которые были установлены на том же слайде изображения с помощью программного обеспечения Leica LCS. Срезы наблюдались также с использованием Nikon ( Мелвилл, Нью-Йорк, США).
Клеточные культуры.
Бигликан-дефицитная линия мышечных клеток была создана с использованием утвержденных протоколов (16). Бигликан нулевые клетки выращивали до ~ 75% слияния на покрытых желатином Permanox chamberslides ( Nalge Nunc, Naperville, Иллинойс, США ) при 33 ° С и 10% углекислого газа в ростовой среде, содержащей модифицированную Dilbecco Eagle ( DMEM) с высоким содержанием глюкозы (КЗС ), 20% FBS, 2% экстрактом куриного эмбриона, 1% L-глутамина, 1% пенициллина / стрептомицина, и 1U INF - г. Клетки дифференцированы 4-9 дней в среде, содержащей DMEM высокий Glc, 5% лошадиную сыворотку, 1% L-глутамин и 1% пенициллин / стрептомицин.
Иммуноцитохимия
Дифференцированные бигликан - нулевые клетки инкубировали в течение 4 ч при 33 ° С с 0,7 нМ. Ацетилхолин (АХ ) рецепторы были помечены родамин - б - бунгаротоксином в течение 30 мин при 33 ° С Дальнейшая инкубация была завершена при комнатной температуре и клетки промывали в MEM - H после каждого шага. Клетки фиксировали в 1 % параформальдегиде и 0,05% сапонина. Для обнаружения нОАС клетки инкубировали с первичном Aт в течение 1 ч, затем инкубируют с козьими анти - мышиными IgG Alexa 488 течение 1 часа. Клетки фиксировали метанолом в течение 5 мин при -20 ° С ( Vector Laboratories, Burlinghame, CA, USA). Изображения были получены на микроскопе Nikon Eclipse E800 с использованием 60. Сегменты без каких-либо поверхностно - локализованных нОАС получили оценку 0 и сегмент с наибольшей степенью получил 4 .
Внутримышечные инъекции бигликан.
Пятьдесят мкг очищенного рекомбинантного бигликана в 50 мкл 20 мМ Трис, 0,5 М NaCl, 0,2% CHAPS, рН 8.0/1.0 % вводили внутримышечно в правую четырехглавую мышцу бедра 2- недельных старых бигликан нулевых мышей с использованием 29 1/2G инсулинового шприца (BD Biosciences, Сан-Хосе, Калифорния, США). Пятьдесят мкл 20 мМ Трис, 0,5 М NaCl, 0,2 % CHAPS, рН 8.0/1.0 % вводили в левую четырехглавую мышцу бедра каждого животного, чтобы служить в качестве внутреннего контроля. Инъецированные четырехглавый мышцы были выделены через 4, 7, 11 и 14 дней после инъекции бигликана. Инъекции у 2 – недельных старых животных были проведены с использованием трех BGN нулевые пометов. Увеличение б - дистробревина -1, б - дистробревина -2 , в1 - синтрофина и в2 - синтрофина через 11 дней после инъекции наблюдалось у 4 из 4 животных и увеличение через 14 дней после инъекции наблюдали в 4 из 4 животных. Никакого увеличения не наблюдалось ни в одной из контрольных мышц. Для иммуногистохимической
![]()
маркировки мышечной ткани, четырехглавые мышцы бедра были выделены как описано выше. Срезы были дважды подвергнуты действию моноклональных антибигликановых антител 2А5 или 4C6 и кроличьих поликлональных антител против указанного дистробревина или синтрофина (см. АнтиВторичные антитела, используемые для обнаружения: Alexa Fluor 488- козы против IgG мыши и Cy-3 козьих антител против кроличьего IgG в разведении 1:2000 .
РЕЗУЛЬТАТЫ
Бигликан-дефицитные мыши обнаруживают дистрофический фенотип.
Мутации в любом из нескольких компонентов DAPC вызвают широкий спектр мышечных дистрофий ( 5 , 17 ). Бигликан связывается с тремя компонентами: б - дистрогликаном, б - саркогликаном и г - саркогликаном (3) (MS Rafii, Х. Хагивара, и JR Fallon, неопубликованные результаты ). Мы проверяли, действительно ли отсутствие бигликана у мышей вызывает дистрофию (рис. 1 ). Во-первых, мы провели анамиз поглощения синего красителя Эванса для определения целостности мембран мышц находится под угрозой. Бигликан нулевым мышам и дистрофин нулевым MDX мышам внутривенно вводили синий краситель Эванса и оценивали степень поглощения в скелетных мышечных волокнах после 6 часов. Мышечные клетки дикого типа имели малую проницаемость (< 0,5% клеток ) , в то время как > 90% MDX мышечных волокон показали поглощение красирис. 1А ) . Бигликан нулевые волокна отображают промежуточное состояние. Одна популяция волокон была полностью проницаема и показала равномерное распределение красителя (7,4% ± 1,3 ; п = 3) , тогда как другие показали околомембранное распределение. Эта локализация на периферии клетки не наблюдалось у контрольных мышц из помета дикого типа. Измерение сывороточных уровней КФК также показало умеренную сарколемную проницаемость у бигликан нулевых мышей. Мы наблюдали уровни 240 ± 43 , 870 ± 104 и 6435 ± 385 для дикого типа, бигликан дефицитных и MDX мышей, соответственно ; U / L, ± SE, п = 3) . Таким образом, субпопуляции мышечных волокон у бигликан нулевых мышей показывают свидетельство дырявых мембран и потери целостности сарколеммы.
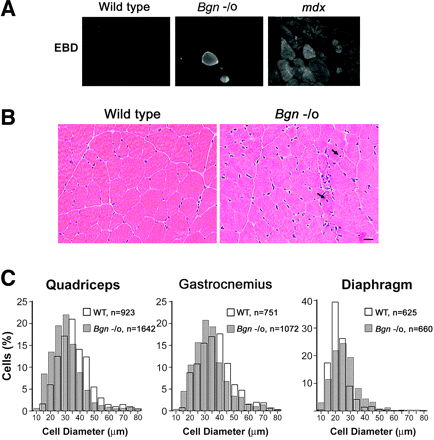
Мы рассмотрели гистологию мышц от бигликан нулевых мышей. Как показано на рис. 1B, большинство волокон в бигликан нулевых мышцах были аналогичны дикому типу, о чем можно судить по окрашиванию гематоксилином и эозином. Тем не менее, доля мышечных волокон у бигликан нулевых мышей имеют центрально локализованные ядра, отличительная черта регенерирующих волокон ( 5.08 % ± 0,58 и 9,65 % ± 1,34 в четырехглавой и диафрагмы, соответственно). В отличие от волокон <0,5% волокон у животных дикого типа имели центральные ядра. Подобный процент ядер в центре волокна были обнаружены в четырехглавой мышцы у 4 - , 12 - и 24- недельных с отсутствием бигликана (данные не показаны). Мы не наблюдали обширную клеточную инфильтрацию мононуклеаров или фиброз у бигликан нулевых мышц. Аномальные мышцы часто характеризуются повышенной изменчивостью размера волокон. Поэтому мы измеряли диаметр
![]()
волокон четырехглавой мышцы бедра, икроножной мышцы и диафрагмы (рис. 1С ). В четырехглавой и икроножной бигликан нулевых мышцах волокна были меньше, чем у дикого типа. Средний диаметр волокна в четырехглавой мышце дикого типа был 36,89 ± 3,88 мкм против 30,56 ± 2,73 мкм в бигликан нулевых мышцах. Средний диаметр волокна в икроножной мышце дикого типа был 38,14 ± 4,69 мкм против 33,67 ± 3,52 мкм при отсутствие бигликана. С другой стороны, диафрагма из бигликан нулевых мышей отображает более широкий гранулометрический состав волокна (фиг. 1C). Средний диаметр размера волокна в мышцах дикого типа диафрагмы был 23,20 ± 3,08 мкм против 27,89 мкм ± 5.21 мкм в диафрагме с отсутствием бигликана. Взятые вместе, эти результаты показывают, что бигликан - нулевые мыши обнаруживают умеренный дистрофический фенотип.
Селективное восстановление б - дистробревина, синтрофина.
Мы сравнили экспрессию отдельных DAPC белков у взрослых дикого типа и бигликан нулевых мышей. Во всех случаях мы сравнивали бигликан нулевых мышей с нормальными или конгенными, подобранными по возрасту животными дикого типа. Результаты были совместимы между животными. Сначала мы исследовали уровни экспрессии дистрофина, основных трансмембранных комплексов, и ламинин б2 . Как показано на рис. 2, иммунофлуоресцентное окрашивание срезов мышц показало, что сарколемная экспрессия дистрофина и б - , в - , и г - саркогликана остается неизменной у бигликан нулевых животных. Экспрессия б - и в - дистрогликана и б2 цепи ламинина были также неразличимы у всех групп мышей (рис. 2).
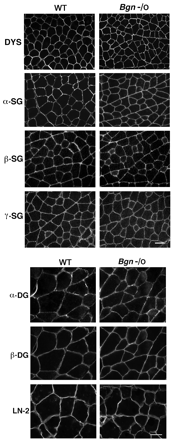
Мы также использовали иммуногистохимию, чтобы изучить экспрессию внутриклеточных DAPC белков, участвующих в передаче сигналов. В каждом эксперименте мы также подвергли ткани иммуноокрашиванию для дистрофина, который является неизменным и таким образом служит в качестве внутреннего, положительного контроля (рис. 2 ⇓ и рис. 3 ⇓ А, В). Как показано на рис. 3A, б-дистробревин-1 и -2 сильно экспрессированы в мышечной сарколемме дикого типа мышц. Кроме того, эти белки локализованы внепрерывно, непрерывно распределены на сарколемме. В противоположность этому, у BGN нулевой мышцы, б-дистробревин-1 и -2 выражены на более низком уровне в сарколемме и распределены нерегулярно, точечным рисунком.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
