
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
![]()
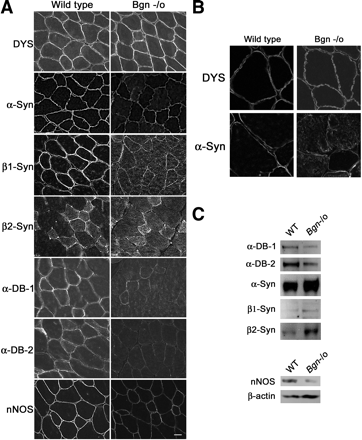
Мы также отметили, что б - синтрофин и в1 - синтрофин снижаются в сарколемме бигликан - дефицитных животных (рис. 3А). С другой стороны, экспрессия в2 - синтрофина в сарколемме бигликан нулевой мышцы не отличалась от мышей дикого типа. Внутриклеточный уровень всех трех синтрофинов был увеличен в бигликан нулевых по сравнению с мышцами дикого типа (рис. 3а, б). Как показано на рис. 3А, сарколемные уровни ННО были ниже в бигликан нулевых мышцах, чем у мышей дикого типа. Взятые вместе, эти результаты показывают, что существует селективное восстановление в экспрессии комплекса дистробревин-синтрофин - нОАС в сарколемме бигликан нулевых мышей. Практически все компоненты DAPC экспрессируются в нервно-мышечном соединении. Кроме того, люди с нарушениями денервации показали специфические изменения в паттернах экспрессии этих синтрофинов и дистробревинов, но не остальную часть DAPC (21). Таким образом, мы спросили, была ли изменена экспрессия б - дистробревина, синтрофина и нОАС в синапсах в бигликан нулевой мышце. Мы считаем, что уровни экспрессии и распределения б - дистробревина -1 , б - дистробревина -2 , б - синтрофина, в1 - синтрофина, в2 - синтрофина и ННО остаются неизменными в бигликан нулевых нервно-мышечных синапсах ( данные не представлены) . Таким образом, этот DAPC подкомплекс уменьшается в сарколемме но поддерживается в нервно-мышечном соединении. Нарушение регуляции б - дистробревина -1 и -2 , б - , в1 - и в2 - синтрофина, и ННО мембранных фракций из бигликан нулевой мышцы. Иммуногистохимические данные, описанные выше, показывают, что уровни б - дистробревина -1 и -2 уменьшены на плазматической мембране бигликан - дефицитных мышечных волокон. Чтобы определить, изменяется ли биохимический профиль этих белков, мы оценили свои уровни в KCl - промытых тяжелых микросомальных мембранных фракций, которые являются смесью плазмы и внутриклеточных мембран (22). Чтобы сравнить уровни мембран-связаного дистробревина, мы выделили фракции от дикого типа и бигликан нулевых четырехглавых мышц и исследовали их с помощью пан - специфических анти-дистробревин антител. Рисунок 3С показывает, что уровни как б - дистробревин -1 и -2 уменьшены в этих мембранных фракциях. Мы рядом исследовали экспрессию б - , в1 - и в2 - синтрофинов в KCl - промытых мембранных фракциях. Иммуноблоттинг с изоформами - специфических антител показал, что уровни всех трех синтрофинов были увеличены в мембранных фракциях при отсутствии бигликана по сравнению с контролем дикого типа (фиг. 3C). Это повышенная экспрессия, вероятно, отражает вклад синтрофинов связанных с внутриклеточными мембранами; действительно, это находится в согласии с нашим иммуногистохимическими результатами показывая повышенные внутриклеточные уровни всех синтрофинов в бигликан нулевых мышцах (рис. 3а, б). Эти результаты показывают, что бигликан служит для
![]()
соответствующего целевого прикрепления синтрофина к поверхности мышечных клеток и / или регулировать внутриклеточный транспорт этих молекул. Мы также сравнили экспрессию нОАС дикого типа и бигликан нулевых. Как показано на рис. 3C, уровни экспрессии нОАС в мембранах бигликан нулевых животных снизилась по сравнению с диким типом, в согласии с иммуногистохимическими результататами. Избирательные изменения в1 - синтрофина и уровней экспрессии мРНК ННО в скелетных мышцах бигликан нулевых мышей.
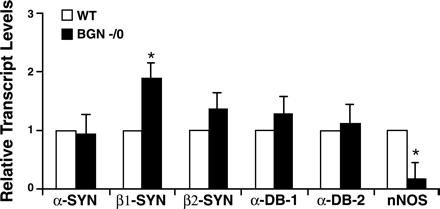
Количественный анализ ПЦР в реальном времени ( QRT - PCR) показал, что транскрипты в1 - синтрофина значительно повышали регуляцию у бигликан нулевых мышей по сравнению с диким типом ( 1,88 ± 0,27 против 1,00 , P < 0,03 , Т критерий Стьюдента ) , в то время уровни транскриптов ННО регулируются по принципу отрицательной обратной связи ( 0,18 ± 0,27 против 1,00 , р <0,03 , Т - критерий Стьюдента ) . С другой стороны, уровни обоих б - дистробревинов и б - и в2 - синтрофинов эквивалентны дикому типу и бигликан нулевым мышцам.
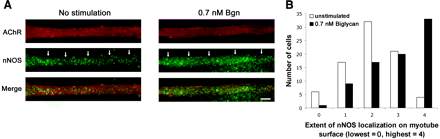
Бигликан вызывает перераспределение нОАС к плазматической мембране.
Мы разработали систему для культивирования клеток для дальнейшего изучения роли бигликана в ориентации DAPC компонентов к поверхности клетки. Мы культивировали бигликан - дефицитные мышечные трубки и инкубировали их либо с очищенным рекомбинантным полипептидом ядром бигликана или только наполнителем в течение 4 часов. Мышечные трубочки были помечены родамин - б - бунгаротоксином для визуализации АХР и разграничения плазматической мембраны. Затем мы фиксировали и подвергли иммунному окрашиванию на наличие дистробревинов, синтрофинов и нОАС. Как показано на рис. 5, ННО распределяется по всей цитоплазме в необработанных клетках, с небольшой маркировкой, наблюдаемой в области плазматической мембраны. Тем не менее, лечение бигликаном повышает уровни локализованного ННО. Мы количественно оценили это перераспределение нОАС путем присвоения каждой ячейке баллов (0-4 0 - отсутствие, 4 самый высокий уровень нОАС на поверхности), представляющих степень присутствующего на поверхности нОАС. При отсутствии бигликана, средний балл составил 2,0 ± 0,11 ; в присутствии бигликана, средний балл 2,94 ± 0,12 . Гистограмма на рис. 5B отображает количество неочищенных или бигликан обработанных клеток. Эти данные показывают, что лечение BGN нулевых мышечных трубок ядром бигликана увеличивает количество нОАС на мембране ( P <0,01 ; тест Колмогорова-Смирнова ).
В параллельных экспериментах мы отметили, что окрашивание б - дистробревина -1 и -2 и в2 - синтрофина присутствует внутри клетки. С другой стороны, были экспрессированы б - синтрофин и в1 - синтрофин по всей клетке, в том числе в поверхностно - проксимальном расположении. Тем не менее, лечение бигликаном не изменяет локализацию этих белков.
Инъекция очищенного бигликана восстанавливает экспрессию б - дистрофинов и в - синтрофинов в сарколемме BGN нулевых мышечных волокон в естественных условиях.
Результаты, описанные выше, показывают, что подкомплекс DAPC уменьшается в сарколемме взрослых BGN нулевых мышей. Однако, поскольку отсутствует бигликан в ходе развития, то невозможно определить, когда бигликан играет роль в локализации этих компонентов. Кроме того, поскольку бигликан выражен в других местах, в том числе костях и сухожилиях ( 23 , 24) , то возможно, что фенотип имеет вторичный дефект в другой ткани. Наконец, результаты у BGN нулевых мышей не позволяют определить, что требуется (ядро или протеогликан ) для направления правильной локализацию DAPC подкомплекса. Очищенный
![]()
рекомбинантный полипептид бигликан, ядро или протеогликан (50 мкг ), инъецировали в правую четырехглавую мышцу бедра 2- WK - старых BGN нулевых животных. Буфер о вводили в левую четырехглавую мышцу, чтобы представить контроль внутри животного. Место инъекции визуализировали путем метки тушью. Четырехглавую мышцу рассекали через 4, 7, 11 и 14 дней после инъекции, делали срезы и проводили иммунологическое исследование.

Внутримышечно вводят ядро бигликан, что оказывает поразительное влияние на экспрессии синтрофинов и дистробревинов в BGN нулевой мышце. К 11 дней после инъекции, мы наблюдаем повышенную экспрессию б - дистробревина -1 и -2 и в1 - и в2 - синтрофина в сарколемме. Кроме того, повышенная экспрессия этих внутриклеточных DAPC белков показало тесную пространственную корреляцию с экзогенным бигликаном (рис. 6а - D). Повышающей регуляции не наблюдалась при введении (рис. 6А - D ) ядра или протеогликанов бигликана ( данные не показаны). Увеличение б - дистробревина -1 и -2 и в1 - и в2 - синтрофина сохранялась на 14 дней после инъекции. Мы не наблюдали изменения в б - синтрофина после инъекции. В самом деле, есть свидетельства, что механическая травма индуцирует экспрессию ННО в мозге (25). Взятые вместе, эти результаты показывают, что бигликан может быть доставлен в мышцы в естественных условиях, и что он может направить локализацию б - дистробревинов и в - синтрофинов к сарколемме. Таким образом, бигликан может регулировать экспрессию комплекса дистробревин-синтрофин на сарколемме в естественных условиях.
ОБСУЖДЕНИЕ
В этом исследовании мы демонстрируем, что BGN нулевые мыши имеют мягкую мышечную дистрофию, и обеспечивают несколько линий доказательств, что это внеклеточный белок регулирует локализацию и экспрессию подмножества внутриклеточных DAPC компонентов в скелетных мышцах. Более того, наши данные показывают, что механизмы, лежащие в бигликанопосредованной экспрессии синтрофинов, дистробревинов и нОАС, вероятно, будут различны. Здесь мы обсуждаем доказательства, подтверждающие эти выводы, потенциальные основные механизмы и последствия этих выводов для потенциальной терапии для мышечной дистрофии. Бигликан-дефицитные мышцы отображают мягкую мышечную дистрофию и характеризуются повышенной КФK сыворотки и поглощения синего красителя Эванса, увеличение числа централизованных ядер в волокнах, и ненормальным распределением размеров миофибрилл. Мембрана была наиболее пострадавшей у ~ 10 % волокон, показывающих центральные ядра. Эти особенности, наблюдаемые на 5 неделе, не были прогрессивными и не сопровождались мононуклеарной клеточной инфильтрацией. Стоит отметить, что этот фенотип качественно подобен тому, что наблюдается у б - дистробревин нулевых мышей ( которые также показывают снижение ННО на сарколемме ; реф 26 ⇓ ) . В этом случае наблюдается ~ 50%
![]()

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
