
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
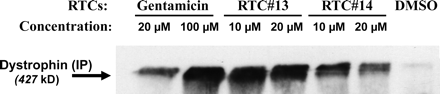
RTC индуцированный MDX PTC в мышечных клетках мыши.
Для исследования возможности RTC № 13 и № 14 исправления мутации в генах, кроме АТМ, мы использовали миофибриллы MDX мышей. Мыши MDX были широко использованы в качестве модели для мышечной дистрофии Дюшенна. Она несет замещение C на T в экзоне 23 гена дистрофина, который создает преждевременный стоп-кодон (ТАА), и в результате отсутствие белка дистрофина (Sicinski и соавт., 1989). Поскольку гентамицин, как было показано, вызывает исправление мутации MDX и восстанавливает экспрессию дистрофина у MDX мышей, как в пробирке и в живом организме (Barton-Davis и соавт., 1999), мы использовали его в качестве положительного контроля в экспериментах. Мышцы клеток-предшественников, выделенных из мышей MDX были вынуждены дифференцировать в течение 24 часов перед добавлением гентамицина или соединения RTC № 13 и № 14. Культуры миофибрилл подвергались обработке соединением в течение 72 часов, а затем проанализированы на наличие дистрофина при помощи вестерн блоттинга. Дистрофин во всю длинну явно обнаруживается в клетках, обработанных RTC # 13 и # 14 в концентрациях от 10 до 20 мкм, но не в культурах, обработанных гентамицином в
![]()
концентрации 20 мкМ (неопубликованные данные). Затем мы использовали иммунопреципитацию для повышения чувствительности анализа (рис. 8). Экспрессия дистрофина присутствует в клетках, обработанных гентамицином в концентрациях от 20 до 100 мкм. Оба наших соединения показали активность, вызывая образование значительного количества белка дистрофина.
Обсуждение
Мы считаем, что HTS может выявить новые неаминогликозидные RTC с терапевтическим потенциалом. В этом исследовании мы использовали в качестве модели заболевания для выявления новых РУЦ. Атаксию-телеангиоэктазию, прогрессивное аутосомно-рецессивное нейродегенеративное расстройство в результате мутации в гене ATM (Перлман и др., 2003;. Чун и Гатти, 2004). АТ белок играет очень важную роль в клеточном цикле, репарации ДНК, окислительном стресс-ответе, и апоптозе (Шило, 2006). В спектре мутаций ATM, основная нонсенс-мутация составляет ~ 15% уникальных мутаций обнаруженных у больных АТ (www. LOVD. nl / ATM). LCLS полученные от пациентов с мутациями, позволяют исследовать влияние неаминогликозидных RTC на различную первичную преждевременную остановку кодонов. Ранее мы уже использовали аминогликозиды и антисмысловые олигонуклеотиды для исправления носенс и мутации сплайсинга ATM, соответственно, для восстановления функциональных белков АТМ в AT LCLS (Lai и др., 2004;.. Ду и др., 2007). Эти исследования показывают, что терапевтический эффект может быть достигнут, если даже незначительное повышение функционального уровня белка могут быть вызваны.
При этом, мы успешно разработали чувствительные люциферазы для независимого анализа HTS по связи PTT и ИФА. PTT-ИФА демонстрирует высокую специфичность для обнаружения продуктов и, таким образом, минимизирует число ложных срабатываний в начальной крупномасштабной части скрининга. Анализ также очень чувствителен, минимальный порог обнаружения составляет ~ 1%, что обеспечивает его эффективность. Эффективность PTT-ИФА в дальнейшем оценивается с помощью двух известных RTC, G418 и гентамицина. Анализ был в состоянии обнаружить их активность в очень больших концентрациях (G418, 40 нм-10 мкл, гентамицин, 40
![]()
нм-100 мкл). Кроме того, РТТ-ИФА была утверждена для полностью автоматизированной платформы. 384-а формат значительно сократит нагрузку на скрининг тысячи соединений, а также экономит время и реагенты. По этим причинам мы считаем, что PTT-ИФА HTS анализ дает новый мощный инструмент для выявления новых RTC.
Из списка ~ 34 000 соединений, мы определили 12 низкомолекулярных соединений массой (от ~ 300 до 450 дальтон), с активностью PTC. Ни одно из этих новых соединений не было аминогликозидом. Несколько соединений показали EC50 значения <10 мкМ, что означает наличие у них потенциала для дальнейшего развития. Для дальнейшей оценки этих веществ в клеточных системах, мы протестировали их деятельность в AT LCLS с различными нонсенс-мутации. Мы использовали ATM-ИФА для непосредственного обнаружения АТМ белка в клетках. Впоследствии, более чувствительные клетки на основе анализов, таких, как FC на основе SMC1-Ser966, ATM-Ser1981 фосфорилирования, и ATM-Ser1981-IRIF, были использованы для оценки восстановления АТМ киназы. Среди 12 соединений, RTC № 13 и № 14 показали, PTC деятельность в клетках, как в LCLS, так и фибробластов, о чем свидетельствует ATM-ИФА, ATM киназы (аутофосфорилирование ATM и транс-фосфорилирования SMC1) и CSA. Для определения того, могут ли эти соединения воздействовать на мутации в других генах, мы выбрали клетки мыши MDX и проверили их способность вызывать схожие эффекты у другого вида, другого типа клеток (неделящихся мышечных клеток), а также различные преждевременные стоп-кодоны (ТАА ). В других исследованиях, TAA кодон оказался самым трудным для прочтения(Kimura и др., 2005;.. Уэлч и др., 2007). Оба RTC # 13 и # 14 индуцировали образование у мыши MDX дистрофина.
В совокупности наши данные показали, что оба RTC # 13 и # 14 были сопоставимы по деятельности на основе анализов, хотя их деятельность в бесклеточных PTT-ИФА были намного ниже, чем у гентамицина и G418. Это может быть связано с различиями в клетках обмена веществ, растворимости и проницаемости для этих соединений.
Для оценки воздействия этих соединений на остановку нормальных кодонов других белков, мы провели двумерный гель-электрофорез. Ни одно из соединений существенно не мешает экспрессии белка (рис. S4), подразумевая, что эти соединения имеют потенциал для дальнейшего развития. Тем не менее, мы ожидаем, что значительный терапевтический эффект, возможно, будет не заметн у AT пациентов, если RTC - индуцированной ATM уровень будет <15% от нормы, а это весьма вероятно, относится и к другим генетическим нарушениям. Таким образом, будет необходимы дальнейшие структурные изменения для улучшения фармакодинамики. Структурная оптимизация улучшит деятельность и снизить токсичность аминогликозидов (Нудельман и др., 2006;.. Rebibo-Саббах и др., 2007).
Основные механизмы деятельности RTC для этих вновь выявленных соединений остаются неизвестными. Было показано, что все известные RTC влияют на рибосомный синтез. Конечно, аминогликозиды взаимодействовуют с декодирующим центром рибосомальной субъединицы 16S и вызвают мисинкорпорацию из аминокислот на сайте РТС, который позволяет продолжить трансляцию (Килинг и Bedwell, 2005; Zingman и др., 2007.). PTC124 Считается также, что они действовуют в другом месте на рибосоме (Linde и Керем, 2008). Будет интересно узнать, взаимодействуют ли RTC # 13 и # 14 с рибосомой.
A-T атаксия-телеангиоэктазия
ATM A-T мутация
CSA оценка жизнеспособности колоний
FC проточная цитометрия
FI интенсивность флюоресценции
HRP пероксидаза хрена
HTS высокочувствительный скрининг
IR ионизирующее излучение
IRIF очаги, вызванные облучением
LCL лимфобластная линия клеток
мРНК матричная РНК
NMD нонсенс опосредованный распад мРНК
PTC преждевременная остановка кодона
PTT транскрипция/ трансляция протеина
RTC соединения для PTC
SMC структура хромосом
Ссылки
↵ Auld, D. S., N. Thorne, W. F. Maguire, J. Inglese. 2009. Mechanism of PTC124 activity in cell-based luciferase assays of nonsense codon suppression. Proc. Natl. Acad. Sci. USA. 106:3585–3590. doi:10.1073/pnas.0813345106 Abstract/FREE Full Text
↵ Barton-Davis, E. R., L. Cordier, D. I. Shoturma, S. E. Leland, H. L. Sweeney. 1999. Aminoglycoside antibiotics restore dystrophin function to skeletal muscles of mdx mice. J. Clin. Invest. 104:375–381. doi:10.1172/JCI7866 Medline
↵ Bedwell, D. M., A. Kaenjak, D. J. Benos, Z. Bebok, J. K. Bubien, J. Hong, A. Tousson, J. P. Clancy, E. J. Sorscher. ppression of a CFTR premature stop mutation in a bronchial epithelial cell line. Nat. Med. 3:1280–1284. doi:10.1038/nm1197-1280 CrossRef
![]()
Medline
↵ Bertoni, C., T. A. Rando. 2002. Dystrophin gene repair in mdx muscle precursor cells in vitro and in vivo mediated by RNA-DNA chimeric oligonucleotides. Hum. Gene Ther. 13:707–718. doi:10.1089/104303402317322276 CrossRef
Medline
↵ Butch, A. W., H. H. Chun, S. A. Nahas, R. A. Gatti. 2004. Immunoassay to measure ataxia-telangiectasia mutated protein in cellular lysates. Clin. Chem. 50:2302–2308. doi:10.1373/clinchem.2004.039461 Abstract/FREE Full Text
↵ Chun, H. H., R. A. Gatti. 2004. Ataxia-telangiectasia, an evolving phenotype. DNA Repair (Amst.). 3:1187–1196. doi:10.1016/j. dnarep.2004.04.010 CrossRef
Medline
↵ Chun, H. H., n, S. A. Nahas, S. Teraoka, C. H. Lai, P. Concannon, R. A. Gatti. 2003. Improved diagnostic testing for ataxia-telangiectasia by immunoblotting of nuclear lysates for ATM protein expression. Mol. Genet. Metab. 80:437–443. doi:10.1016/j. ymgme.2003.09.008 CrossRef
Medline
↵ Chun, H. H., R. B. Cary, F. Lansigan, J. Whitelegge, D. J. Rawlings, R. A. Gatti. 2004. ATM protein purified from vaccinia virus expression system: DNA binding requirements for kinase activation. Biochem. Biophys. mun. 322:74–81. doi:10.1016/j. bbrc.2004.07.085 CrossRef
Medline
↵ Clancy, J. P., Z. Bebцk, F. Ruiz, C. King, J. Jones, L. Walker, H. Greer, J. Hong, L. Wing, M. Macaluso, et al. 2001. Evidence that systemic gentamicin suppresses premature stop mutations in patients with cystic fibrosis. Am. J. Respir. Crit. Care Med. 163:1683–1692. Abstract/FREE Full Text

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
