
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Неаминогликозидный состав устраняет носенс мутации

Неаминогликозидный состав устраняет носенс мутации.
Liutao Du1, Robert Damoiseaux5, Shareef Nahas1, Kun Gao2, Hailiang Hu1, Julianne M. Pollard1, Jimena Goldstine3, Michael E. Jung6, Susanne M. Henning2, Carmen Bertoni4, and Richard A. Gatti1,3
Author Affiliations
1Department of Pathology and Laboratory Medicine, 2Center for Human Nutrition, 3Department of Human Genetics, and 4Department of Neurology, David Geffen School of Medicine, 5Molecular Shared Screening Resources, California NanoSystems Institute, 6Department of Chemistry and Biochemistry, University of California, Los Angeles, Los Angeles, CA 90095
Author Notes
L. Du and R. Damoiseaux contributed equally to this paper.
CORRESPONDENCE Richard A. Gatti: *****@***ucla. edu OR Liutao Du: *****@***ucla. edu
Введение
Большое количество генетических нарушений вызваны носенс мутациями, для которых состав, индуцирующий преждевременную остановку кодонов (PTC), может быть использован в качестве потенциальной стратегии лечения. Мы успешно разработали чувствительный и количественный высокопроизводительный скрининг (HTS) транскрипции / трансляции (PTT) белка, иммуноферментный анализ (ИФА) для выявления новых PTC - соединений с использованием атаксии-телеангиэктазии (AT), как генетической модели заболевания. Это HTS PTT-ИФА на основе PTT который использует шаблоны, содержащие плазмиды прототипных AT (ATM) мутаций для HTS. Мы изучили ~ 34 000 соединений и определили 12 низкомолекулярных неаимногликозидных соединения с потенциальной PTC активностью. Из них два ведущих соединения последовательно индуцирующих функциональные белки в ATM-дефицитных клетках, содержащих болезнетворные нонсенс-мутации. Прямые измерения ATM белка показали восстановление ATM киназы. Эти два соединения также продемонстрировали активность в миофибриллах MDX мышей, несущих носенс мутации, и индуцированных значительное количество белка дистрофина.
Об окончании трансляции сигнализируют три кодона: UAA, UAG и UGA. Этот механизм хорошо сохранилися, хотя каждый кодон имеет различную эффективность для завершения трансляции. UGA считается "дырявым" стоп-кодоном с высоким внутренним потенциалом для остановки кодона. UAA показывает высокую точность и малый внутренний потенциал, в то время как UAG имеет промежуточные значения (Weiner и Вебер, 1973. Ловетт и др., 1991). Нонсенс-мутации создают первичную преждевременную остановку кодонов (PTC) и в результате либо не образуется белок или синтезируется усеченный белк с нарушением стабильности.
Некоторые соединения влияют на точность остановки кодона и вызывают первичную PTC, которая позволяет транскрипцию некоторых полных белков. Во многих случаях, вызванный этим событием белок функционален, даже если он содержит ошибочно включенные аминокислоты (Килинг и Bedwell, 2005;. Zingman и др., 2007).
Считается, что 30% человеческих болезнетворных аллелей – это нонсенс-мутации (Менделл и Dietz, 2001). Другие типы мутаций, таких, как смещение рамки считывания и мутации сплайсинга, приводят к вторичной PTC, однако, это не может быть мишенью для терапевтических целей. Учитывая, что> 1800 различных генетических нарушений вызвано нонсенс-мутациями, первичная PTC имеет потенциал для лечения большого количества больных.
На сегодняшний день большинство из соединений, вызывающих PTC, которые активны в клетках млекопитающих, принадлежали к классу аминогликозидов (Килинг и Bedwell, 2005;. Zingman и др., 2007). Некоторые виды аминогликозидов могут помочь рибосомам прочитать мутации через вставки случайной аминокислоты. Терапевтический потенциал аминогликозидов был оценен в лаборатории для различных генетических моделей, таких как кистозный фиброз (Howard и др., 1996;. Bedwell и др., 1997;.. Ду и др., 2002), мышечная дистрофия (Howard и др.. , 2000; Вагнера и др., 2001;. Дюнан и др., 2003;. Loufrani и др., 2004), Hurler синдром (Килинг и др., 2001), цистиноз (Helip-Вули и др., 2002), спинальная мышечная... атрофия (Sossi и др., 2001)., атаксия-телеангиэктазии (AT,. Лай и др., 2004), и тип 1 синдрома Ушера (Rebibo-Саббах и др., 2007.). Клинические исследования также показывают, что аминогликозиды могут вызывать некоторое производство функционального белка, но терапевтический эффект остается неопределенным (Wilschanski и др., 2000;. Clancy и др., 2001;. Вагнер и др., 2001;.. Politano и др., 2003).
Кроме того, токсичность большинства коммерческих аминогликозидам у млекопитающих значительно снижает их потенциал для успешной терапии (Mingeot-Леклерк и Тюлькенс, 1999; Гуан и др., 2000.). Таким образом, предпринимаются усилия по разработке производных аминогликозидов с пониженной токсичностью и повышенной активностью (Нудельман и др., 2006;.. Rebibo-Саббах и др., 2007). В последнее время PTC Therapeutics (Южная Plainfield, Нью-Джерси), описала более эффективное неаминогликозидное соединение PTC124, которое было разработано синтетическим путем скрининга> 800 000 химических веществ и аналогов с помощью люциферазы на основе высокопроизводительного скрининга (HTS) (Welch и соавт., 2007 ; Hirawat и др., 2007;. М. Ду и др., 2008)..Фазы-I клинических исследований при муковисцидозе подтвердила, что PTC124, как правило, хорошо переносится и, как представляется, более эффективен, чем аминогликозиды (Hirawat и соавт., 2007). Кроме того, PTC124 не вызывает рибосомных эффектов, нормализует остановку кодонов. Фаза-II клинических испытаний ведется в настоящее время (Керем и соавт., 2008). Однако, недавнее исследование показывает, что начальные открытие PTC124 может быть предвзятым.
В попытке открыть для себя новые соединения, мы разработали чувствительный и количественный независимый анализ HTS транскрипции / трансляции белка (PTT). PTT-ИФА была разработана для автоматизированной 384-а платформы и используется для скрининга ~ 34 000 соединений. Мы сосредоточили усилия на последующих 12 низкомолекулярных неаминогликозидных соединенях. Оттуда, мы определили два соединения, индуцирующих низкий уровень полного функционального белка во всю длинну в клетках, несущих AT нонсенс-мутации, как показали прямые измерения ATM белка с использованием ATM-ИФА, ATM-Ser1981 аутофосфорилирования, транс-фосфорилирования структур хромосомы (SMC) 1-Ser966 и анализа выживания колонии. Оба соединения также показали активность в отношении миофибрилл MDX мышей, несущих носенс-мутации, и индуцировали значительное количество белка дистрофина.
В совокупности эти исследования дают первые надежные подтверждения надежности независимого HTS анализа для выявления таких соединений и доказывают принцип PTC для неаминогликозидных соединений. Они также устанавливают, что расширение возможностей PTC можно рассматривать как терапевтическую стратегию для исправления нонсенс-мутации при многих генетических заболеваниях.
Результаты
Разработка и утверждение HTS анализа
In vitro PTT был первоначально разработан для обнаружения мутаций (Roest и др., 1993;.. Telatar и др., 1996). ИФА использовалось в сочетании PTT чтобы улучшить пропускную способность для выявления этих мутаций (Gite и др., 2003;.. L. Du и др., 2008). В предыдущем исследовании мы использовали PTT - в геле для оценки различных аминогликозидов по воздействию на нонсенс-мутации (Lai и соавт., 2004). Тем не менее, анализа PTT в геле занимал много времени и связан с использованием радиоактивных материалов, поэтому было сложно автоматизировать его для высокой пропускной способности. При этом, мы разработали плазмиды для PTT-ИФА для скрининга большого числа соединений PTC. Анализ использует шаблоны, содержащие плазмиды прототипных мутаций AT, по образцу конкретных болезнетворных мутаций AT. Так как требовалась работа в клетках млекопитающих, были выбраны ретикулоциты кролика для управления реакцией PTT. Различные фрагменты мутировавших аллелей из клеток больных АТ были клонированы в плазмиды с N-и С-терминальной меткой эпитопов myc и V5, соответственно. Anti - myc антитела использовались для получения перевода белка на пластины ELISA. Если соединения вызывают PTC в анализе, в полнометражных ATM фрагмент включался V5, которая обнаруживается антителами анти-V5-пероксидазой хрена (HRP) (рис. 1).
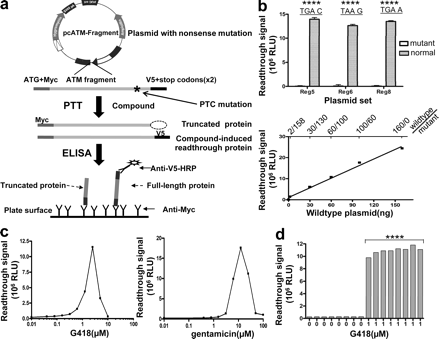
Мы построили три мутантные плазмиды, содержащие прототипные мутации в АТ гене от трех разных больных АТ. Плазмиды используются для предварительного отбора, плазмиды TAT51, содержит ATM области 5 фрагмент (кодонов 1403-1886) и таит в себе носенс - мутацию (c.5623C → T), что приводит к остановке кодона TGA. Вторая плазмиды AT153LA содержит тот же стоп-кодон TGA, но на другой позиции внутри гена (c.8977C → T) в области 8 (кодонов 2550-3050). Она была использована для мониторинга влияния окружающей последовательности на PTC
![]()
способности. Третья мутантная плазмида AT185LA, содержит различные стоп-кодоны, TAA G, в результате нонсенс мутации (c.3673C → T) в области 4 (кодонов 1041-1531). Плазмиды, содержащие те же фрагменты, но без мутаций были построены в пробирке мутагенезом производных комплементарной ДНК пациентов и были использованы в качестве дикого типа контроля.
Сначала мы оценивали специфичность и чувствительность PTT-ИФА для определения в пробирке трансляции полнометражного фрагмента белка. Все три мутантные плазмиды дали только фоновый сигнал, в то время как сопоставимые плазмиды дикого типа показали сигналы больщее ~ 200 раз по сравнению с фоном (рис. 1, б, вверху), что указывает на специальность фрагментов белка. Затем мы использовали TAT51 плазмиду дикого типа для оценки чувствительности анализа. TAT51 дикий тип плазмиды серийно разбавляют TAT51 мутантной плазмидой и используется для управления PTT реакцией. Образцы, содержащеие 1,2% дикого типа плазмид (2/158 нг) дали сигнал, который был вдвое выше, чем у мутантных плазмид (рис. 1, б, внизу), установив чувствительность теста ~ 1%.Чувствительность анализа HTS особенно важная для скрининга РTC, потому что все RTC на сегодняшний день были только слабыми PTC индукторами, и новые классы RTC должны быть определены высоко чувствительным скринингом.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
