

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
6 | 7 | 8 | 9 | 10 | |
A | 0.2±0.1 1.0±0.2↑ | 0.4±0.2 1.5±0.2↑ | 0.4±0.2 1.3±0.1↑ | 0.2±0.1 0.4±0.1↑ | |
B | 0.9±0.3 2.0±0.2↑ | 1.9±0.5 2.8±0.2↑ | 2.2±0.4 2.4±0.2 | 1.1±0.4 1.6±0.2 | 0.4±0.3 0.4±0.1 |
C | 2.2±0.5 2.5±0.2 | 4.8±0.9 3.1±0.2 | 5.8±0.8 3.0±0.2↓ | 2.3±0.5 2.2±0.1 | 0.6±0.2 1.1±0.1↑ |
D | 5.2±0.9 2.2±0.2↓ | 5.6±0.8 2.8±0.1↓ | 5.1±0.6 2.9±0.1↓ | 2.2±0.3 2.6±0.2 | 0.5±0.2 1.7±0.3↑ |
E | 3.0±0.3 2.4±0.2 | 3.2±0.4 2.9±0.1 | 1.4±0.3 2.8±0.2↑ | 1.7±0.4 2.5±0.1↑ | 0.3±0.1 0.9±0.2↑ |
Примечание: в числителе дано распределение при плотности заселения листа 1 мина на лист, в знаменамин на лист. Достоверные различия (Р<0.05) обозначены стрелками: ↑ - в случае повышения, ↓ - в случае понижения показателя. Во всех случаях n=10.
Таким образом, распределение мин липовой моли-пестрянки в нижнем ярусе кроны кормового растения носит равномерный характер независимо от экспозиции ветви и плотности заселения растения. Бабочки предпочитают откладывать яйца в зоне развитого жилкования – базальной и средней частях листа. Увеличение плотности L. issikii до 10 мин на лист приводит к равномерному распределению мин по листу липы сердцевидной.
Глава 8. Динамика численности липовой моли-пестрянки
и факторы ее определяющие
Состояние популяции характеризуют такие показатели, как соотношение полов, изменчивость окраски, размеров особей, плодовитость, деятельность паразитических насекомых и другие (Справочник…, 1980).
Стационарные исследования динамики численности липовой моли-пестрянки на трех пробных площадях показали, что колебания численности в популяциях минера происходят на фоне относительно стабильного полового индекса (Табл. 7.). Сохранение оптимального соотношения полов является важнейшим механизмом поддержания генетической разнородности популяций (Шварц, 1980, с. 161).
Таблица 7.
Динамика полового индекса первого поколения липовой моли-пестрянки (Малиновая гора)
Год | N1 | Плотность заселения, мин на 100 листьев | N2 | Самцы, % | Самки, % |
2001 | 40 | 155.5±14.8 | 2253 | 52.1±1.1 | 47.9±1.1 |
2002 | 40 | 120.6±12.3 | 1214 | 47.6±1.4 A | 52.4±1.4 A |
2003 | 40 | 169.0±14.2 | 1946 | 51.9±1.1 B | 48.1±1.1 B |
2004 | 40 | 897.7±41.5 | 1516 | 46.8±1.3 C | 53.2±1.3 C |
2005 | 40 | 478.7±29.7 | 1644 | 53.3±1.2 D | 46.7±1.2 D |
Примечание: N1 – количество модельных деревьев; N2 – количество исследованных бабочек; достоверные отличия (Р<0,05) отмечены одинаковыми буквами
Согласно работе Т. Кумата (Kumata, 1963а), бабочки липовой моли-пестрянки характеризуются сезонным диморфизмом: представители летней морфы имеют золотисто-охристую, осенней - серую окраску крыльев. Собственные исследования показали, что оба варианта, “темный” и “светлый”, встречаются в каждой генерации (Табл. 8.) Отношение между ними широко варьирует в зависимости от плотности.
Таблица 8.
Отношение между морфами бабочек первого поколения липовой моли-пестрянки, % (Малиновая гора)
Год | N1 | N2 | Плотность заселения, мин на 100 листьев | Самцы | Самки | ||
“темные” | “светлые” | “темные” | “светлые” | ||||
2002 | 40 | 1214 | 120.6±12.3 | 15.2±1.0 | 32.4±1.4 | 23.3±1.2 | 29.1±1.3 |
2003 | 40 | 1946 | 169.0±14.2 | 45.9±1.1 | 6.0±1.7 | 43.8±1.1 | 4.3±1.4 |
2004 | 40 | 1516 | 897.7±41.5 | 44.9±1.3 | 2.0±1.1 | 51.3±1.3 | 1.8±1.1 |
2005 | 40 | 1644 | 478.7±29.7 | 50.9±1.2 | 2.4±1.1 | 44.3±1.2 | 2.4±1.3 |
Примечание: N1 – количество модельных деревьев; N2 – количество исследованных бабочек
Морфометрический анализ бабочек показал, что длина крыла со “светлым” вариантом окраски достоверно меньше аналогичного показателя крыла “темной” окраски (Табл. 9.).
Таблица 9.
Длина левого переднего крыла бабочек липовой моли-пестрянки, мм (Малиновая гора)
Год | N1 | Самцы | Самки | ||
“темные” | “светлые” | “темные” | “светлые” | ||
2002 | 39 | 3.25±0.01* (N2=185) | 3.24±0.01** (N2=393) | 3.19±0.01* H (N2=283) | 3.15±0.01** H (N2=353) |
2003 | 40 | 3.18±0.01* (N2=893) | 3.16±0.02** (N2=117) | 3.11±0.01* I (N2=853) | 3.05±0.02** I (N2=83) |
2004 | 40 | 3.21±0.01*A (N2=680) | 3.10±0.04 A (N2=30) | 3.13±0.01* J (N2=778) | 2.99±0.05 J (N2=28) |
2005 | 40 | 3.30±0.01*B (N2=837) | 3.09±0.04 B (N2=39) | 3.21±0.01* K (N2=728) | 3.13±0.03 K (N2=40) |
Примечание: N1 – количество модельных деревьев. Достоверные различия (P<0.05) между морфами одного пола отмечены одинаковыми буквами, между полами одной морфы – звездочками (* - для “темных”, ** - для “светлых”).
Изменение плотности заселения липы L. issikii сопровождается трансформацией структуры популяции минера (Рис. 4.). Повышение плотности заселения модельных деревьев приводит к снижению доли “светлых” бабочек на всех пробных площадях: “Малиновая гора” (r=-0.49; P<0.001; n=160), “Питомник” (r=-0.42; P<0.001; n=157) и “Парк им. ” (r=-0.60; P<0.001; n=160).
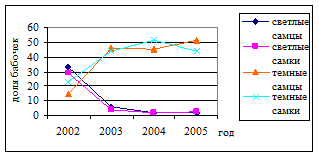
Рис. 4. Влияние плотности заселения деревьев минером на соотношение бабочек разных морф (Малиновая гора). Примечание: по горизонтали – год исследования, по вертикали – доля бабочек (%).
Исследование содержимого яичников самок L. issikii в период откладки яиц показало (Табл. 10.), что самки с “темными” крыльями достоверно (Р<0.001) более плодовиты, чем самки со “светлым” вариантом крыла.
Таблица 10.
Основные характеристики бабочек-самок липовой моли-пестрянки, собранных 25.05.04 на десяти стволах липы мелколистной
Самки | Длина, мм | Количество яиц в яичниках, шт. | ||
тела | крыла | |||
“темные” | 142 | 3.55± 0.02 А | 3.23±0.01 В | 32.4±1.0 С |
“светлые” | 28 | 3.26±0.04 А | 3.04±0.03 В | 23.7±1.4 С |
Примечание: N – количество исследованных бабочек; достоверные различия (Р<0.001) отмечены одинаковыми буквами.
Влияние факторов, ограничивающих численность минера, изучено в течение 2002 – 2005 гг. Данные по выживаемости L. issikii на стадии куколки приведены в таблице 11. Результаты исследований, проведенных на трех пробных площадях, показали, что популяции моли имеют крайне низкую степень регуляции со стороны паразитов (от 0.4±0.1 (n=40) до 13.5±1.5% (n=38)). Для сравнения, смертность от паразитизма в популяциях L. populifoliella и L. crataegella Clemens достигала 74% (Румянцев, 1934) и 76,7% (Driesche, Taub, 1983) соответственно.
Значительно большее количество куколок липовой моли-пестрянки (до 34.3±2.0% (n=40)) погибает в минах в результате действия неизвестных причин (заражение патогенами, неудачные попытки выхода бабочек из куколок, антибиотическая реакция дерева, деятельность куколочных паразитов). В результате выживаемость генераций моли превышает 60%.
Таблица 11.
Характеристики выживаемости и смертности куколок L. issikii (Малиновая гора)
Год | N | Плотность заселения, мин на 100 листьев | Выживаемость, % | Смертность, % | |
Паразитизм | Неизвестная причина | ||||
2002 | 40 | 120.6±12.2 | 64.1±2.0 | 1.6±0.4 | 34.3±2.0 |
2003 | 40 | 169.0±14.2 | 70.2±1.3 | 1.2±0.3 | 28.6±1.3 |
2004 | 40 | 897.7±41.5 | 70.5±1.6 | 0.4±0.1 | 29.1±1.7 |
2005 | 40 | 478.7±29.7 | 79.6±1.7 | 0.5±0.2 | 19.9±1.7 |
Примечание: N – количество модельных деревьев.
Таким образом, колебания численности в популяциях липовой моли-пестрянки в диапазоне от 17.5 до 897.7 мин на лист происходят на фоне стабильного полового индекса. Для бабочек L. issikii характерен полиморфизм по окраске крыла. Увеличение плотности заселения растения-хозяина приводит к повышению доли “темных” бабочек, достоверно отличающихся большей длиной крыла и плодовитостью. Выживаемость генераций моли превышает показатель в 60%.
Выводы
1. Липовая моль-пестрянка широко распространена по всей территории Удмуртии и может давать два поколения.
2. Взаимоотношения в системе “липа - липовая моль-пестрянка” носят активный характер. Увеличение плотности заселения липы минером приводит к увеличению площади мины.
3. Минер способен заселять липовые насаждения вне зависимости от их возраста, физиологического состояния и происхождения. Пространственное распределение мин первого поколения моли по скелетным ветвям нижнего яруса носит равномерный характер освоения вне зависимости от экспозиции ветви и плотности заселения растения. В условиях низких плотностей заселения выявлены участки наибольшего предпочтения при яйцекладке моли. При повышении плотности происходит выравнивание распределения мин по листу. Количество мин на листе положительно и достоверно связано с его площадью.
4. Изменение численности в популяциях липовой моли-пестрянки в диапазоне от 17.5 до 897.7 мин на 100 листьев происходит на фоне стабильного полового индекса. Вспышки массового размножения минера являются следствием слабых регулирующих механизмов. Смертность от паразитов колеблется в пределах между 0.4±0.1 (n=40) и 13.5±1.5% (n=38), гибель от неизвестных причин составляет от 14.7±1.1 (n=40) до 34.3±2.0 (n=40) %. В результате выживаемость генераций моли составляет значительные величины (от 64.1±2.0 (n=40) до 81.7±0.9% (n=40)). Увеличение плотности заселения растения-хозяина приводит к повышению доли “темных” морф бабочек, достоверно отличающихся большей длиной крыла и плодовитостью. Подобные изменения, вероятно, оказывают влияние на пассивное расселение вида.
Список работ, опубликованных по теме диссертации:
1. , Мотошкова (Ижболдина) моль-пестрянка Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) в Удмуртии // Тезисы докладов XII съезда РЭО, Санкт-Петербург,августа 2002 г. - Санкт-Петербург: ЗИН РАНС.
2. , Мотошкова (Ижболдина) влияния липовой моли-пестрянки на линейный прирост побегов дерева-хозяина // Труды региональной научно-практической конференции “Аграрная наука – состояние и проблемы”. - Том II. - Ижевск: ИжГСХАС.
3. , Мотошкова (Ижболдина) липовой моли-пестрянки на линейный прирост побегов дерева-хозяина // Биоразнообразие и биоресурсы Урала и сопредельных территорий. Материалы II Международной конференции. Оренбург, 17-18 декабря 2002 г. - Оренбург: ОГПУС.
4. , Мотошкова (Ижболдина) Н. В. О биологии липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Разнообразие беспозвоночных животных на Севере: Тезисы докладов II Международной конференции (Сыктывкар, Республика Коми, Россия, 17-22 марта 2003 г.). - Сыктывкар: Коми НЦ УО РАНС.
5. , Мотошкова (Ижболдина) моль-пестрянка – новый экономически значимый филлофаг липовых лесов Удмуртии // Структурно-функциональная организация и динамика лесов. Материалы Всероссийской конференции. - Красноярск: ИЛ СО РАНС.
6. , Мотошкова (Ижболдина) влияния липовой моли-пестрянки на прирост и генеративную сферу липы мелколистной // Методы популяционной биологии. Сборник материалов VII Всероссийского популяционного семинара. - Ч.1. - Сыктывкар: ИБ КНЦ УрО РАНС.
7. , Мотошкова (Ижболдина) биологии липовой моли-пестрянки в Удмуртии // Биоразнообразие природных и антропогенных экосистем. - Екатеринбург: УрО РАНС.
8. , Мотошкова (Ижболдина) Н. В. О полиморфизме в популяциях липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Лесоводство и лесные мелиорации. Материалы межвузовской научно-практической конференции. - Новочеркасск: НГМАС.
9. , Мотошкова (Ижболдина) в популяциях липовой моли-пестрянки // Материалы VII научно-практической конференции преподавателей и сотрудников УдГУ, посвященной 245-летию г. Ижевска. Часть 2. - Ижевск: УдГУС.
10. , Мотошкова (Ижболдина) полиморфизма в популяциях липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Принципы и способы сохранения биоразнообразия. Сборник материалов II Всероссийской научной конференции. - Йошкар-Ола: МГУС.
11. , Мотошкова (Ижболдина) освоения липовой молью-пестрянкой Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) липы мелколистной // Биоразнообразие и биоресурсы Урала и сопредельных территорий. Материалы III международной конференции. - Оренбург: Принт-сервисС.
12. , Мотошкова (Ижболдина) инвазия на примере липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Энтомологические исследования в Северной Азии. Материалы VII Межрегионального совещания энтомологов Сибири и Дальнего Востока (в рамках Сибирской зоологической конференции). - Новосибирск: ИСиЭЖ СО РАНС.
13. , Мотошкова (Ижболдина) полиморфизма в популяциях липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) в биологической инвазии вида // Особь и популяция - стратегии жизни. Материалы IX Всероссийского популяционного семинара. - Ч. 2. - Уфа: Вили ОкслерС.
14. , Мотошкова (Ижболдина) повреждения листьев липы мелколистной липовой молью-пестрянкой. Гипотеза природных маркеров // Международная научная конференция “75 лет высшему образованию в Удмуртии”: Материалы конференции: Ч. 2. Естественные науки. - Ижевск: УдГУС.
15. , Мотошкова (Ижболдина) выживаемости генераций липовой моли-пестрянки // Инновационное развитие АПК. Итоги и перспективы: материалы Всероссийской научно-практической конференции. - Т.1. - Ижевск: ФГОУ ВПО ИжГСХАС.
16. , Мотошкова (Ижболдина) моль-пестрянка // Защита и карантин растений№ 5. - С.
17. , Мотошкова (Ижболдина) биологической инвазии липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Синантропизация растений и животных. Материалы Всероссийской конференции с международным участием. - Иркутск: ИГ СО РАНС.
18. , Мотошкова (Ижболдина) Н. В. О механизмах биологической инвазии липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Достижения энтомологии на службе агропромышленного комплекса, лесного хозяйства и медицины. Тезисы докладов XIII съезда Русского энтомологического общества, Краснодар, 9-15 сентября 2007 г. - Краснодар: КГАУС.
19. , Мотошкова (Ижболдина) инвазия липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae): особенности взаимоотношения минера с кормовым растением // Энтомологическое обозрениеТ. 87. - № 1. - C.
20. , К механизмам инвазии липовой моли-пестрянки Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae): оценка выживаемости генераций минера // XXII Любищевские чтения. Современные проблемы эволюции. Том 2. - Ульяновск: УГПУС.
21. , Ижболдина инвазии липовой моли-пестрянки в Европе // Современное состояние и пути развития популяционной биологии. Материалы Х Всероссийского популяционного семинара (17-22 ноября 2008 г.). - Ижевск: Книгоград, 2008. - С.
22. , Зорин распределения мин липовой моли-пестрянки в системе: древостой – дерево – лист // Известия Санкт-Петербургской лесотехнической академииВып. 182. - С.
23. Ermolaev I. V., Motoshkova (Izhboldina) N. V. Biology of leaf miner Lithocolletis issikii Kumata (Lepidoptera, Gracillariidae) // Invertebrate animals diversity in the North: Abstracts of the Second International Conference. (Syktyvkar, the Komi Republic, Russia, March 17-22, 2003). - Syktyvkar, 2003. - P.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
