

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
10. Значение сорных растений в формировании структуры сообщества почвенной мезофауны
10.1 Структура сообщества почвенной мезофауны
В зернопаровом севообороте и на поле с бессменной пшеницей, на взаимное распределение различных групп почвенных беспозвоночных влияет антропогенный фактор нарушенности естественных сообществ, но в разных сорных растительных ассоциациях такое влияние ощущается по-разному. Для проверки этого допущения исследовали характер корреляционных связей между обилием разных групп мезофауны в сорных растительных ассоциациях.
На учетных участках почвы с минимальными и нулевыми зяблевыми обработками в сорных растительных ассоциациях Galeopsetum bifidae и Cannabio-Sinapetum установлена положительная связь между численностью пауков и другими массовыми группами почвенной мезофауны. Обратная зависимость не установлена между массовыми группами почвенной мезофауны.
В сорных растительных ассоциациях Lathyro-Lactucetum tataricae, Lactucetum-tataricae, Lactuco-Artemisietum и Lactuco-Carduetum при переходе к минимальным обработкам почвы, на участках даже в условиях полной химической нагрузки (УФГ), проявляется положительный характер связей между численностью массовых групп почвенной мезофауны. Негативный эффект антропогенных нагрузок в этих сорных растительных сообществах сглаживается и менее всего отражается на беспозвоночных обитающих на поверхности почвы и в верхнем замульчированном слое. Для тлей, клопов, многих перепончатокрылых, двукрылых и златоглазок в данных сорных растительных сообществах численность хотя и имеет положительную зависимость, но не всегда является достоверной.
10.2 Сезонный порядок распределения беспозвоночной мезофауны
На примере сообщества жужелиц, установлено, что итоговый набор обитаемых участков с различными зяблевыми обработками и особенно количественное предпочтение каких-либо из них, является у большинства видов (но не у всех без исключения) следствием непрерывного процесса «подстраивания» к сезонной динамике экологической обстановки в зернопаровом севообороте и на бессменной пшенице, путем соответствующего пространственного комбинирования в пределах определенного растительного сорного сообщества популяционным фондом особей беспозвоночных. Предпочитаемыми участками вида будут те, где экологические условия оптимальны для вида в течение наиболее длительного времени. Так, например, последовательная смена предпочитаемых участков на озимой ржи и на пшенице - второй культуре после пара, в течение сезона хорошо выражена у видов Amara bifrons, A. apricaria, Microlestes minutulus в сорных растительных ассоциациях Galeopsetum bifidae. Эти виды, например, в мае 2001 года при среднемесячной температуре воздуха +16,1ºС (при выпавших 16,3 мм осадков против среднемноголетних 36 мм), наиболее многочисленные видовые группировки имели на вспашке; в июне со среднемесячной температурой (+17,7ºС), близкой к среднемноголетней (+16,7ºС), но с обильными осадками (87,5 мм против 58 мм среднемноголетних осадков) – на минимальной; в холодном (+16,7ºС) и переувлажненном (115,6 мм) июле – на нулевой; а с августа – на участках с глубокой безотвальной обработкой, когда влажность почвы сохранялась на уровне 16-17%, а плотность – 1,07 г/см3 и оставалась в таком режиме в сентябре. Как видим, сезонные смены предпочтения носят довольно резкий характер. Альтернативные смены предпочтения обитания в агрофитоценозе обычно характерны для видов с невысокой плотностью особей в популяции (Harpalus smaragdinus, Ophonus puncticollis). У видов богатых особями смены носят количественный характер медленного постепенного «переползания» (Мордкович, 1985) ядра популяции на участки агрофитоценоза с более благоприятными условиями обитания. Так у вида M. minutulus, несмотря на смещение в мае ядра популяции от вспашки на участки с минимальной зяблевой обработкой почвы, крупная видовая группировка осталась на вспашке.
10.3 Топическое распределение видов беспозвоночных в зависимости от многолетней динамики экологического режима в агрофитоценозе
На примере с жужелицами доказано, что агрофитоценоз с зернопаровым севооборотом и вне его - на поле с бессменной пшеницей представляет возможность виду (и беспозвоночных животных и сорных растений) в ходе сезонной и многолетней динамики популяций реализовать весь запас своих адаптивных потенций. Агрофитоценоз с его определенной структурой антропогенного фона предопределяет неравномерное размещение популяционного фонда особей вида вдоль градиента условий предпочтительной и менее предпочтительных почвенных режимов, что и позволяет определить агроэкологический оптимум и пределы отклонения от него, а затем описать и сравнить по единой шкале экологические стандарты множества живых взаимосвязанных и взаимозависимых организмов, в том числе, сорных растений и беспозвоночных животных.
10.4 Трофические взаимоотношения и связи с сорными растениями
В результате изучения трофических связей на примере подземногнездящихся пчел рофитоидес канус, мелитта лепорина и мелиттурга клавикорнис в условиях зернового агрофитоценоза получено убедительное доказательство относительного постоянства трех подземногнездящихся видов диких пчелиных к собиранию пыльцы с сорных растений из семейства бобовых, причем предпочтение отдается пыльце люцерны посевной.
Установлено, что три вида подземногнездящихся пчел во все годы исследований выступают единым опылительным комплексом, т. е. все три вида пчел обязательно присутствуют одновременно на общем кормовом участке зернового агрофитоценоза, причем присутствие рофитоидеса кануса является показателем наличия нектара на данном участке для мелитты лепорины и мелиттурги клавикорнис. Положительная корреляционная связь фуражирующих самок между парами видов рофитоидес канус и мелитта лепорина, рофитоидес канус и мелиттурга клавикорнис подтверждается с достоверностью в 98,7% и 99,4% соответственно. Такая положительная взаимосвязь видов подземногнездящихся опылителей на кормовом участке объясняется значительной разницей (более 30%) в длине хоботка в каждой паре, различным механизмом вскрытия цветка.
10.5 Конкурентные взаимоотношения беспозвоночных
на кормовом участке
Для проверки гипотезы об опылительном комплексе видов подземногнездящихся пчел и о сигнальном значении самок рофитоидес канус для остальных насекомых-опылителей сорного сообщества в зерновом агрофитоценозе была проведена серия экспериментов по искусственному изъятию самок рофитоидес канус с кормового участка. Выяснилось, что после изъятия большей части фуражирующих самок рофитоидес канус, плотность мелиттурги клавикорнис и мелитты лепорины уменьшилась почти в два раза (от 1,2 до 0,5 шт./м2). В процессе восстановления численности рофитоидеса кануса, через три дня, плотность мелиттурги клавикорнис и мелитты лепорины снова нарастает до прежнего соотношения.
Установлено, что между показателями фуражировочной активности мелитты лепорины и медоносной пчелой существует отрицательная линейная зависимость:
У=11,6+(-0,953)×Х, (10.5.1)
где RS= -0,651; Р=99,4%;
Отрицательная линейная зависимость существует и между показателями фуражировочной активности медоносной пчелы и мелиттурги клавикорнис, при которой значительное увеличение плотности подземногнездящихся пчел вызывает незначительное уменьшение плотности медоносной пчелы на общем кормовом участке зернового агрофитоценоза:
У=1,433+(-0,061)×Х, (10.5.2)
где RS = 0,426; Р=95,9%;
Корреляция плотности самок рофитоидес канус с плотностью медоносной пчелы на фуражировочном участке недостоверна, т. е. находится ниже принятого уровня значимости.
Таким образом, способность подземногнездящихся пчелиных поселяться в зерновых агрофитоценозах большими скоплениями - агрегациями, обеспечивает устойчивый постоянный комплекс опылителей, в котором доминирующую и определяющую роль играет пчела рофитоидес канус. Конкуренция трех подземногнездящихся видов на кормовом участке ослаблена в результате приобретения ими морфофизиологических дифференцировок, позволивших занять различные экологические ниши. Медоносная пчела выступает в роли конкурента за пищевые ресурсы в зерновом агрофитоценозе для подземногнездящихся пчел
10.6 Хищнические и паразитические взаимоотношения
Условно связанных с подземногнездящимися пчелами животных можно объединить в три группы; хищников, объектом которых являются имаго пчел на время их полетов, паразитов, развивающихся как в имаго, так и в расплоде, и комменсалов-животных, не наносящих ущерба подземногнездящимся пчелам.
Из насекомых в зерновых агрофитоценозах на фуражирующих самок подземногнездящихся пчел нападают крупные ктыри рода Laptria Mg. и некоторые паукообразные. Гибель пчел рофитоидес канус от нападений хищников составляет в среднем 3,3+1,5%.
Наиболее ощутимый вред приносят муравьи рода Lasius L., моментально поселяясь в колониях – гнездовьях непосредственно в зерновом агрофитоценозе. Совершая регулярные набеги в неглубокие гнезда пчелы мелиттурги клавикорнис, могут уничтожить в среднем до 35,7 + 6,3% отрытых гнезд.
Из насекомых, паразитирующих на имаго в районе наших исследований, следует выделить мух рода Conops L. Самка мухи атакует чаще всего мелиттургу клавикорнис в полете и прикрепляет яйцо между стернитами брюшка. В большом количестве они отмечались на участках с вариантами зяблевой вспашки, что приводило к заражению за сезон в среднем 7,1 + 2,5% самок подземногнездящихся пчел.
Из перепончатокрылых - паразитов подземногнездящихся пчел необходимо назвать ос - сапигид из рода Sapyga L. Пораженность ячеек сапигидами составляла в среднем 12,3 + 4,5 от общего числа ячеек обследованных гнезд. Сходно поступает и другой паразит подземногнездящихся пчел из рода Coelioxys L. (семейство Megachilidae). Разница лишь в том, что этот паразит может откладывать яйца в любое время, вплоть до появления личиночной стадии хозяина-пчелы. Поражаемость ячеек этим паразитом в среднем несколько выше и достигала 15,7 + 5,6%.
Как клептопаразиты подземногнездящихся пчел нами также отмечены жуки - майки из рода Meloe L. Взрослые жуки питаются на цветах, где откладывают яйца. Число ячеек пораженных жуками-майками относительно высоко и составляла в среднем 9,6 ± 3,3%.
10.7. Смертность
Для выяснения основных причин, оказывающих влияние на формирование и успешное развитие мезофауны зерновых агроценозов на примере агрегации подземногнездящихся пчел, мы сопоставляли результаты визуальных учетов норковых отверстий пчел с данными по абсолютному учету численности методом Джоли-Зебера. Для анализа состояния агрегации подземногнездящихся пчел использовали девять показателей, характеризующих развитие сообщества в зерновом агрофитоценозе.
Регрессионный анализ показывает, что связь численности пчел и коэффициента их воспроизводства может быть описана уравнением:
Х = 3,968-0,5 × С ± 1,97 (10.7.1)
Где Х – коэффициент воспроизводства;
С – численность.
Поскольку показатели являются средними характеристиками, ошибка уравнения регрессии здесь и ниже определялась как ошибка выборочных групп. Зависимость эта обратно пропорциональна и близка к линейной (достоверность линейности 96,04%).
Связь между численностью и показателем 4 выражается уравнением:
X = 12,816 + 0,330 × С ± 6,663, (10.7.2)
где X - процент гибели самок подземногнездящихся пчел вне гнезда до закладки ячеек. Эта связь прямо пропорциональна и также близка к линейной (достоверность 96,66%).
Из анализа факторов влияющих на состояние численности агрегации подземногнездящихся пчел нами был сознательно исключен климатический фактор. Хотя его влияние бесспорно, он по своей сути является фактором внешним и стохастическим и, следовательно не может оказывать регулирующего действия на численность агрегации подземногнездящихся пчел (Гиммельфарб, Гинзбург и др., 1974). Если такие исключить антропогенный фактор, оказывающий наиболее сильное внешнее влияние на численность агрегации подземногнездящихся пчел, то наши данные позволят предположить, что численность агрегации подземногнездящихся пчел регулируется внутренними механизмами, из которых выделяется два фактора: коэффициент воспроизводства самок и смертность самок подземногнездящихся пчел вне гнезда до периода закладки расплодных ячеек.
11. Влияние антропогенного фактора на распределение беспозвоночного населения почвы в сорных сообществах растений
11.1 Изменение почвенной наннофауны
Установлено, что массовые заболевания растений яровой пшеницы, вызываемые бактериальными возбудителями, вследствие минимизации зяблевых обработок – явление довольно редкое, а ухудшение фитосанитарного состояния яровой пшеницы можно компенсировать подбором системы предпосевной подготовки почвы и посева (Синещеков, Краснопёров, Васильева, Щукин, Мухин, 2003). Минимизация зяблевой обработки почвы, и особенно сведение её к нулю, приводило к увеличению распространенности болезни растений пшеницы на 5-8%, и увеличению её индекса в 1.3-1.4 раза по сравнению с контролем. Улучшение фитосанитарной ситуации в посевах пшеницы в фазу всходов при использовании сеялки СЗП-3.6 на безотвальных фонах было отмечено при применении лущильника ЛД-10, а на вариантах со вспашкой – после ранневесеннего боронования ЗБСС-1.Оптимизация структуры и влажности пахотного слоя почвы в ранневесенний период позволяет создавать необходимые предпосылки для получения стабильного урожая при отказе от глубоких зяблевых обработок почвы или сведения их до минимальных.
11.2 Снижение почвенной микрофауны
Тромбидиформные клещи являются постоянными обитателями почвы агрофитоценоза и поэтому велика их роль как индикаторов почвенной среды и её изменения под влиянием технологических приемов по возделыванию зерновых культур. В агрофитоценозе тромбидиформные клещи остаются одной из немногих массовых групп, так как подавляющая часть других микроартропод гибнет при распахивании почвы (Гатилова, 1969; Вайнштейн, 1972; Краснопёров, Слесарев, Синещеков, 2004).
Установлено, что имеет место неуклонная тенденция накопления численности и биомассы тромбидиформных клещей в течение ротации севооборота (табл. 7).
Таблица 7 – Численность (тыс. шт./м2) (А) и биомасса (мг/м2) (Б) тромбидиформных клещей в зависимости от места в севообороте, гг.
Слой, см | Пар | Озимая рожь- 1-я культура после пара | Пшеница- 2-я культура после пара | Пшеница- 3-я культура после пара | Бессменная пшеница | |||||
А | Б | А | Б | А | Б | А | Б | А | Б | |
0-5 | 0,15 | 0,32 | 0,68 | 0,71 | 1,08 | 0,95 | 1,65 | 3,56 | 5,57 | 8,64 |
5-10 | 0,07 | 0,16 | 0,30 | 0,33 | 0,72 | 0,55 | 0,91 | 1,73 | 1,86 | 2,06 |
10-20 | 0,03 | 0,09 | 0,16 | 0,13 | 0,29 | 0,23 | 0,55 | 0,74 | 0,92 | 1,05 |
20-30 | 0,03 | 0,07 | 0,12 | 0,11 | 0,20 | 0,21 | 0,43 | 0,57 | 0,76 | 0,84 |
30-40 | 0,01 | 0,02 | 0,04 | 0,05 | 0,13 | 0,09 | 0,19 | 0,22 | 0,33 | 0,38 |
0-40 | 0,29 | 0,66 | 1,30 | 1,33 | 2,42 | 2,03 | 3,73 | 6,82 | 9,18 | 13,51 |
Максимального значения численность клещей достигает на бессменной пшенице, что связано с накоплением других групп беспозвоночных и большего видового состава сорных растений, которыми питаются и на которых поселяются почвенные клещи. Глубина почвы, на которой предпочитает обитать более 60 % от общей численности тромбидиформных клещей не превышает 5 см от поверхности почвы. Тромбидиформные клещи избегают глубоких отвальных и безотвальных обработок почвы, предпочитая поселяться на минимальных и нулевых обработках. На этих обработках почвы в большинстве сорных растительных ассоциациях прослеживается тенденция к нарастанию численности почвенных клещей.
Тромбидиформные клещи очень чутко реагируют на применение химических средств защиты основной культуры. Наибольшая гибель почвенных клещей происходит в сорных растительных ассоциациях Cannabio-Sinapetum, Cirsio-Potentilletum и Matricario-Taraxacetum при инсектицидных обработках (до 50 – 65 %) (рис. 8).
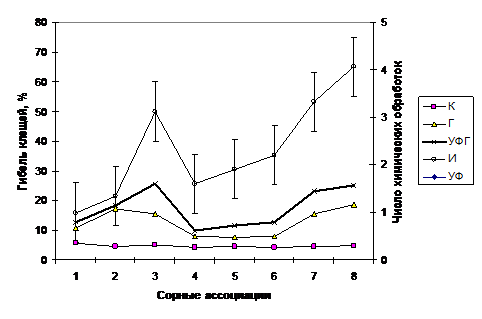
Рис. 8 – Гибель тромбидиформных клещей (%), в зависимости от вида пестицидной нагрузки и количества химических обработок. К – контроль (без химических обработок); Г–гербициды; УФГ–удобрения+фунгициды+гербициды; УФ–удобрения+фунгициды; И–инсектициды.
Это объясняется тем, что названные растительные ассоциации преимущественно произрастают в годы с умеренным переувлажнением и большая часть населения тромбидиформных клещей активна именно на поверхности почвы.
Сокращению численности почвенной микрофауны, в том числе и тромбидиформных клещей способствуют переуплотнение подпахотного слоя, а также непосредственное физическое раздавливание при прохождении сельскохозяйственной техники и прицепных агрегатов. По нашим данным, в весенний период, когда проход по полю агрегатированной техники производится многократно, численность тромбидиформных клещей после трехкратного прохода достоверно сокращалась к моменту посева основной культуры на глубине до 10 см более чем в 2 раза.
11.3 Адаптация надпочвенной мезофауны
Поскольку почвенная мезофауна представляет собой более подвижную структуру зернового агрофитоценоза, беспозвоночные этой группы активнее всего реагируют на изменяющиеся условия обитания, то закономерности их ответной реакции яснее просматриваются на фоне сорных растительных сообществ. Если расположить сорные растительные ассоциации по общему числу обитающих в них видов почвенной мезофауны, получим следующую последовательность: Galeopsetum bifidae; Cannabio-Sinapetum; Lactucetum-tataricae; Lathyro-Lactucetum tataricae; Lactuco-Artemisietum; Lactuco-Carduetum; Cirsio-Potentilletum; Matricario-Taraxacetum. Наименьшее число видов почвенных беспозвоночных животных обитает в сорной ассоциации Galeopsetum bifidae на глубоких безотвальных обработках почвы. Наибольшее число видов почвенной мезофауны - обитателей ассоциации Matricario-Taraxacetum, предпочитают участки агрофитоценоза без зяблевой обработки почвы (нулевая) (рис. 9).
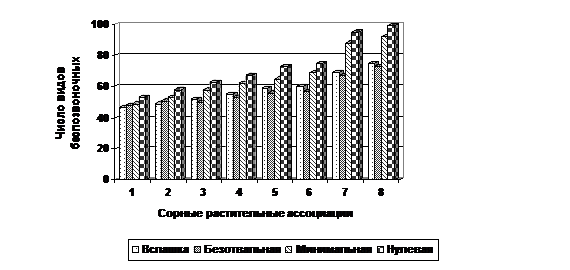
Рис. 9 – Видовое разнообразие мезофауны в сорных растительных ассоциациях в зависимости от системы основной обработки почвы: 1 - Galeopsetum bifidae;2 - Cannabio-Sinapetum; 3 - Lactucetum-tataricae; 4 - Lathyro-Lactucetum tataricae; 5 - Lactuco-Artemisietum; 6 - Lactuco-Carduetum; 7 - Cirsio-Potentilletum; 8 - Matricario-Taraxacetum.
Только в растительных ассоциациях Galeopsetum bifidae и Cannabio-Sinapetum, фаунистическое разнообразие и общая численность беспозвоночных превышают аналогичные показатели на участках с глубокой безотвальной обработкой почвы по сравнению со вспашкой. Во всех остальных сорных растительных ассоциациях фаунистическое разнообразие и общая численность беспозвоночных на вспашке выше, чем при глубоких безотвальных обработках.
При использовании химических средств защиты растений, являющейся составной частью технологических приемов при возделывании яровой пшеницы, то здесь тенденция увеличения видового состава мезофауны отмечена по мере снижения пестицидной нагрузки и максимального значения достигает на участках без зяблевой обработки и без пестицидов (К). Очень близкие значения по видовому разнообразию мезофауны отмечены на участках без зяблевой обработки, но с применением фунгицидов и минеральных удобрений (УФ) (рис. 10).
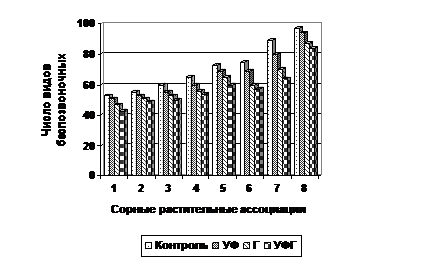
Рис. 10 – Видовое разнообразие мезофауны в сорных растительных ассоциациях в зависимости от пестицидной нагрузки: Контроль – без пестицидов и минеральных удобрений; УФ – фунгициды+минеральные удобрения; Г – гербициды; УФГ – удобрения+фунгициды +гербициды. Сорные растительные ассоциации те же, что на рис. 10
Установлено, что на участках почвы со вспашкой в верхнемсм) слое почвы в среднем обитает до 95 % населения почвенной мезофауны. Крупные беспозвоночные более равномерно распределяются по верхнему см ) слою почвы по мере минимизации обработки почвы, и на участках с систематическим отсутствием зяблевых обработок (нулевая), в верхнем ( 0-10 см ) слое почвы обитает до 85 % от общей численности мезофауны. Глубина заселения основной массы почвенной мезофауны выщелоченного чернозема в зерновом агрофитоценозе совпадает с нижней границей гумусового горизонта.
Значительную долю почвенной мезофауны составляют жужелицы. Доля жужелиц в сообществе герпетобинтов составляла 89 % в фазу кущения пшеницы, снижаясь до 16 % в фазу выхода в трубку. Приемы зяблевой обработки почвы, также как применение средств химической защиты растений оказывали достоверное влияние на большую часть сообщества жужелиц.
По мере удаления яровой пшеницы в системе севооборота от пара и минимизации зяблевой обработки почвы происходит увеличение видового разнообразия сообщества жужелиц.
11.4 Реконструкция подземных гнезд пчел
Более трех четвертей от общего времени активной жизнедеятельности самки подземногнездящихся пчел проводят за устройством гнезда, занимаясь отрытием подземных структур и откладкой яиц. Так для самок рофитоидеса, рабочее время, при посещении сорных растений, составляет в сутки в среднем 126 ± 18 минут, для самок мелиттурги 218 ± 26 минут и для самок мелитты – 268 ± 28 минут. Все остальное время самки пчел проводят в гнезде. Известно, что подземные структуры пчелиных гнезд одного вида различаются от типа грунта (Бондарчук, Радченко, 1985). Однако сведения о том, как адаптируются пчелы к условиям интенсивной антропогенной нагрузки отсутствуют. Нам удалось с помощью оригинальных методик выделить некоторые особенности и различия в гнездостроительной активности пчел внутри агрегации в условиях зернового агрофитоценоза. Для каждого подземногнездящегося вида пчел характерна своя глубина и структура почвенных ходов, поэтому каким образом изменяются их архитектура и какие имеются особенности в условиях зернового агрофитоценоза – эти закономерности помогут определить допустимую степень антропогенной нагрузки и спрогнозировать поселение целого комплекса перепончатокрылых беспозвоночных.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
