

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Гнездовые постройки самок вида рофитоидес канус (рофитоидес). При детальном анализе подземных гнездовых структур выделятся четыре обособленные подгруппы, отличные друг от друга по ряду дополнительных признаков (табл.8).
Таблица 8 – Показатели признаков для четырех подгрупп структуры подземных гнезд Rhophitoides canus Eversm.
Признаки | I группа | II группа | ||
Подгруппы | ||||
1 | 2 | 3 | 4 | |
Среднее отклонение от входа в гнездо (угол, градус) | 7,6 | 31,9 | 57,3 | 25,7 |
На глубине (см) | 8,5 | 10,1 | 1,2 | 6,8 |
Среднее отклонение от входа в гнездо (расстояние, мм) | 0,7 | 4,9 | 1,3 | 2,6 |
На глубине, (см) | 7,4 | 14,8 | 10,9 | 5,1 |
Максимальная длина центрального хода, (см) | 19,4 | 30,7 | 16,7 | 21,4 |
Средний угол при нахождении фронтальной проекции,(градус) | 154,0 | 120,0 | 295,0 | 338,0 |
Максимальное число ячеек в гнезде, (шт.) | 3,0 | 10,0 | 5,0 | 8,0 |
Число раскопанных гнезд | 96 | 226 | 115 | 145 |
Глубина, на которую отрывают свои гнезда самки, в среднем составляет 14,6±1.8 см от поверхности почвы. Максимальная глубина на которой был зафиксирован гнездовой ход – 18.5 см. Боковые ходы заваливается самкой после постройки личиночных ячеек и откладки яиц. Ячеек в боковом ходу обычно 2-3 (очень редко до 5), сферообразной формы - 4,9±0.8 мм в диаметре. Стенки ячеек выкладываются из почвы, смоченной выделениями слюнных желез пчелы и затем шлифуется настолько тщательно, что их внутренняя поверхность выглядит абсолютно гладкой и блестящей. В ячейке находится круглый комочек пыльцы, смешанной со слюной и нектаром, средний вес которого составляет 22,5±2,7 мг. Личинка, закончившая питание, окукливается и кокон заполняет всё пространство ячейки. Стенки кокона состоят из четырёх слоёв: внутреннего шелковистого, слоя экскрементов, воскового и наружного - внутренней почвенной облицовки (Благовещенская, 1982). Этот наружный почвенный слой при раскопке гнезд почти неотделим от кокона.
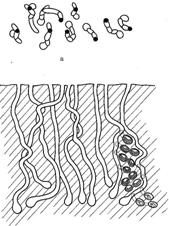
В зерновом агрофитоценозе, например, на фрагменте почвы площадью 10 см2 в одной из агрегаций пчелы рофитоидес, было зафиксировано 9 гнездовых ходов, вырытых за один летний период, но самками разных функциональных групп (рис.11), где проекции центральных ходов на горизонтальную плоскость отображает множество пересечений (см. рис.11, б). В реальности же центральные ходы разнесены в пространстве на 3-5 мм и никогда друг с другом не пересекаются (см. рис.11, а).
а
 б
б
Рис. 11. Схематическое изображение проекций подземных гнезд Rhophitoides canus Eversm. на фрагменте почвы: а) вид сверху; б) вид сбоку (расплодные ячейки даны только для двух гнезд)
Структура подземных гнезд, отрываемых самками мелиттурги также различна, где выделяются два основных типа.
Первый тип – центральный вертикальный вход, самки пчел второй группы, отрывают на небольшую глубину, строят широкую камеру, от которой в противоположные концы ведут коридоры, один из которых тупиковый. Он не ведет к расплодным ячейкам. Другой коридор, более длинный, тянется в горизонтальном направлении и приводит к яйцевым ячейкам (рис. 12, а). Второй тип – самки второй группы строят центральный ход полого уходящий вглубь, без широкой камеры и расходящихся в противоположные стороны коридоров (рис.12 б).
.
а |
б |
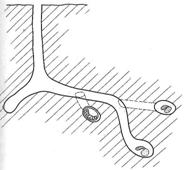
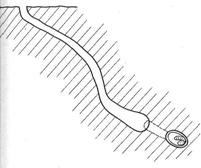
Рис. 12. Характерная архитектура подземного гнезда Melitturga clavicornis Latr. первой (а) и второй группы (б).
Гнезда мелиттурги в агрегациях, чаще всего, располагаются более разреженно в сравнении с пчелами рофитоидес. Так, например, во фрагменте почвы площадью 10 см2содержится 3 гнезда мелиттурги. Гнезда вырыты самками из разных функциональных групп и их проекции не имеют точек соприкосновения друг с другом (рис. 13).
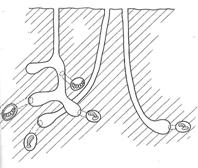
Рис.13. Схематическое изображение проекций подземных гнезд пчелы Melitturga clavicornis Latr. разных групп на фрагменте почвы.
Гнездовые постройки самок пчелы мелитта лепорина. Самки мелитты из основных двух групп строят гнезда также различающиеся по ряду признаков (табл.9).
Таблица 9 - Среднестатистические показатели признаков двух групп структуры подземных гнезд пчелы Melitta leporina Pans.
Признаки | I группа | II группа | Среднее |
Средняя длина центрального хода (см) | 21,2 | 33,6 | 27,4 |
Среднее число ячеек в гнезде | 4,8 | 3,9 | 4,4 |
Средняя глубина, с которой начинаются ячейки, (см) | 6,0 | 16,0 | 11,0 |
Средняя глубина, с которой заканчиваются ячейки, (см) | 11,5 | 18,5 | 15,0 |
Среднее отклонение от входа в гнездо (угол, градус) | 79,0 | 36,0 | 57,5 |
На глубине (см) | 6,0 | 1,0 | 3,5 |
Среднее отклонение от входа в гнездо (расстояние, мм) | 10,0 | 5,6 | 7,8 |
На глубине, (см) | 6,0 | 18,0 | 12,0 |
Средний угол при нахождении фронтальной проекции, (градус) | 65,0 | 270,0 | 167,5 |
Доля законченных гнезд, (%) | 62,0 | 19,5 | 40,7 |
Число раскопанных гнезд | 198 | 215 | ∑=417 |
Из всех описанных выше видов подземногнездящихся пчел наиболее разреженными в условиях зернового агрофитоценоза характерны для мелитты лепорины. На рисунке 14 представлено схематическое изображение подземных построек самок из разных функциональных групп.
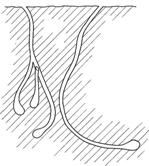
Рис.14. Схематическое изображение проекций подземных гнезд пчелы Melitta leporina Pans. разных групп на фрагменте почвы
Таким образом, подземногнездящиеся пчелы, имеющие тенденцию к поселению агрегациями, способны выдерживать высокую антропогенную нагрузку, поселяясь в условиях зернового агрофитоценоза. Адаптация к интенсивной обработке почвы и применению пестицидов выражается в различии времени постройки гнезда по типу и приему отрывания. Гнезда самок в агрегации могут располагаться чрезвычайно близко друг к другу, но их подземные ходы не пересекаются и не переходят друг в друга. Самки пчел из разных функциональных групп отрывают характерные по конструктивным особенностям гнезда. Это указывает о высокой внутривидовой дифференциации и адаптации к высоким антропогенным нагрузкам в условиях зернового агрофитоценоза.
12. Динамика почвенной мезофауны
Значительный период жизни подземногнездящиеся пчелы проводят в почве зернового агрофитоценоза или тесно соприкасаясь с ним. Поскольку любой агроценоз оказывает на население почвенной мезофауны существенное антропогенное воздействие, а полезное участие этих насекомых в процессе гумусообразования основываются на эксплуатации их естественных популяций, то изучение законов развития и прогнозирование состояний их популяций во времени имеет большое прикладное значение. Экспериментальное изучение многих моментов динамики сообществ подземногнездящихся пчел затруднено, а зачастую и практически невозможно. Такие сложности связаны с рядом биологических особенностей подземногнездящихся пчел: сравнительно низкой плотностью их популяций, достаточно высокой подвижностью преимущественно в дневное время, затрудненность в поиске гнездовий, яйцекладок и т. д. В связи с этим, мы полагаем, целесообразно на основе доступного материала построить общую математическую модель динамики сообщества почвенной мезофауны на примере подземногнездящихся пчел и согласовать ее впоследствии с экспериментальными данными.
12.1 Построение имитационной модели динамики почвенной мезофауны на примере сообщества подземногнездящихся пчел
Для анализа состояния сообщества использовались следующие показатели:1. Численность; 2. Смертность от естественных причин; 3. Смертность от механических повреждений с/х техники; 4. Гибель от других причин; 5. Процент самок, погибших до откладки яиц; 6. Коэффициент воспроизводства.
Колебания численности в сообществах имеют вид осциллирующей кривой (рис.15). При этом резкий спад численности между 1996 и 1997 г. в поле с бессменной пшеницей и между 2000 и 2001 г. в зернопаровом севообороте связан, по нашему мнению, с процессами конформации сообщества, обусловленными на участках с нулевыми зяблевыми обработками эффекта «охранного режима» (режим заповедника), связанного с отсутствием механических обработок почвы. Колебания численности кроме этих лет имеют в обоих случаях синхронный характер, что подтверждает высказанные выше соображения об уровне организации сообщества.
В дальнейшей работе был проведен анализ влияния показателей на численность. При этом данные за 1995-96 гг. по в полю с бессменной пшеницей и за 2000 г. по зернопаровому севообороту были выбракованы как артефакт.
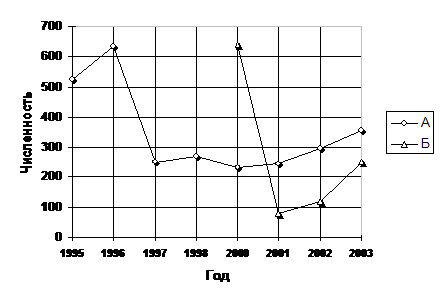
Рис. 15-Динамика численности сообществ подземногнездящихся пчел в поле с бессменной пшеницей (А) и зернопаровом севообороте (Б).
Наибольшая корреляция наблюдалась между численностью и коэффициентом воспроизводства (признак 6) и между численностью и количеством самок, погибших до откладки яиц (признак 5), причем эта закономерность сохранялась как при парном, так и при множественном корреляционном анализе. Коэффициент ранговой корреляции между признаком 1 и 5 составил 0.994 (достоверность 99.98%) а между признаком 1 и 6 - -0,891 (достоверность 99.95%). Корреляция остальных признаков с численностью находилась ниже 89% и не признана достоверной на принятом нами уровне значимости. Результаты множественного корреляционного анализа совпадают с этим результатом в таблице 8. Поэтому в дальнейшем мы ограничились анализом влияния на численность только этих двух факторов.
Таблица 10 - Множественный корреляционный анализ влияния показателей динамики сообщества подземногнездящихся пчел на его численность
Признак | Частный коэффициент корреляции | Достоверность (%) |
2 | 0.428 | 79.6 |
3 | -0,269 | 52.38 |
4 | 0.239 | 47.41 |
5 | -0.123 | 99.07 |
6 | 0.321 | 97.58 |
Коэфф. множественной корреляции | 0.970 | 99.81 |
Регрессионный анализ показывает, что связь численности и признака 6 может быть описана уравнением
X = 12.,040×N ± 1.
где X - коэффициент воспроизводства; N - численность. Поскольку показатели являются средними характеристиками, ошибка уравнения регрессии здесь и ниже определялась как ошибка выборочной группы. Зависимость эта обратно пропорциональна и близка к линейной (достоверность линейности 96.04%).
Связь между численностью и признаком 5 выражается уравнением
X = 12.,040×N ± 1.
где X - процент гибели самок до воспроизводства. Эта связь прямо пропорциональна и также близка к линейной (достоверность линейности 98.66%).
Эти данные позволяют предположить, что численность сообщества подземногнездящихся пчел регулируется, главным образом, внутренними механизмами и в дальнейших расчетах ограничиться рассмотрением этих механизмов. Из дальнейшего рассмотрения также сознательно был исключен климатический фактор. Хотя его влияние на любые организмы бесспорен, он по своей сути является фактором внешним и стохастическим и, следовательно, не может оказывать регулирующего действия на численность сообщества (Гиммельфарб, Гинзбург, Полуэктов, 1974). Поэтому введение его в модель будет лишь помехой для понимания механизмов динамики.
12.2 Имитационная модель динамики сообщества подземногнездящихся пчел
Модель строилась на экспериментальном материале и состояла из имитационного стохастического воспроизведения двух основных процессов динамики – перемещении самок пчел в пределах агрофитоценоза и откладка яиц на участках с нулевой зяблевой обработкой и конкурентного размножения видов пчел в разных сорных растительных сообществах. Модель конкурентного размножения организована по принципу игры «Жизнь», предложенной Дж. Конвэем (Гроссман, Тернер Дж., 1983), где репродукция каждой особи определяется количеством ее соседей. В отличие от детерминированного характера, предложенного Конвэем алгоритма, мы использовали его стохастический аналог. Модель реализована в программе ВОMBUS.
12.3 Результаты моделирования динамики сообщества
подземногнездящихся пчел
Первоначально численность моделируемого сообщества дает в течение 2-х – 3-х лет нерегулярные колебания, обусловленные приведением первоначальной конфигурации сообщества в соответствие с предложенным агрофитоценозом и привязкой к конкретным экологическим условиям предложенной местности. Затем сообщество приходит к некоторому стационарному уровню, как по численности, так и по пространственной конфигурации, и численность его начинает осциллировать с периодом 2 года, подобно реально наблюдавшимся процессам в сообществах подземногнездящихся пчел. Такая динамика находит себе объяснение в триггер-эффекте, характерном для популяций с синхронизированными циклами размножения (Скарлецкая, Фрисман, Шапиро, 1979).
Таким образом, экспериментальные данные по зерновому агрофитоценозу при почвозащитных приемах зяблевой обработки почвы и построенная на их основе имитационная модель представляют сообщество почвенной мезофауны на примере подземногнездящихся пчел как гибкую саморегулирующуюся систему.
выводы
1. При переходе к экологическому адаптивно-ландшафтному земледелию на выщелоченных черноземах лесостепи Приобья в зернопаровом севообороте и на бессменной пшенице в течение нескольких ротаций формируется как минимум восемь сорных растительных сообществ - Galeopsetum bifidae, Cannabio-Sinapetum, Lactucetum-tataricae, Lathyro-Lactucetum tataricae, Lactuco-Artemisietum, Lactuco-Carduetum, Cirsio-Potentilletum и Matricario-Taraxacetum. При сокращении глубины зяблевых обработок до нуля все большее значение будут приобретать ассоциации сорных растений: Латуко-Полынная (Lactuco-Artemisietum), Латуко-Чертополоховая (Lactuco-Carduetum), Бодяково-Лапчатковая (Cirsio-Potentilletum), Трехреберниково-Одуванчиковая (Matricario-Taraxacetum officinalis).
2. В ассоциациях сорных растений Lactuco-Artemisietum, Lactuco-Carduetum, Cirsio-Potentilletum и Matricario-Taraxacetum officinalis более всего представлены розеточные растения (36.0%), меньше всего полурозеточных (30.3%), промежуточное положение занимают безрозеточные растения (33.7%), что хорошо согласуется с установленными закономерностями развития безрозеточных и розеточных растений и их тесной взаимосвязи с экологическими условиями среды. Более прогрессивный, с точки зрения развития растения, розеточный тип побегообразования может противостоять наиболее небагоприятным условиям внешней среды. Подавляющее большинство видов сорных растений являются симподиальными, на долю моноподиальных приходится только 4.5% от всего видового разнообразия.
3. По характеру перезимовки в ассоциациях сорных растений союза Achillion millefolii летнезеленые сорные растения составляют 64.1%, зимнезеленые - 27%, вечнозеленые – 8.9%. Количество летнезеленых сорных растений уменьшается по мере минимизации обработок почвы, что связано с большими конкурентными способностями зимнезеленых и вечнозеленых сорняков. Вечнозеленые растения чаще всего укоренялись на вариантах с нулевой зяблевой обработкой и полностью отсутствовали на минимальной обработке

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
