
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
К 15-18 м характер растительности полностью меняется. Начинают доминировать Сodium vermilara (1,2-2,2 кг/м2; 10-70 экз/м2) и Phyllophora crispa ( г/м2; 20-50 экз/м2), образуя ассоциации Codietum phyllophorosum и Phyllophoretum purum. Цистозира, хотя и присутствует в сообществе, не доминирует и не достигает сколько-нибудь значительного развития. Видовой состав второго и третьего ярусов значительно обеднен: присутствуют Nereia filiformis с биомассой от 30 до 320 г/м2, Apoglossum ruscifolium, Ceramium sp., Chaetomorpha sp., Gracillaria gracilis, Stilophora rhizodes, Jania rubens, Zanardinia prototypus, Polysiphonia subulifera, Corallina elongata, C. granifera, Gelidium latifolium, G. spinosum. Четвертый ярус представлен корковыми водорослями Hydrolithon farinosum (= Melobesia farinosa) и Hildenbrandtia rubra. Общая биомасса фитоценоза не превышает 2500 г/м2, ОПП – 25-65%.
На глубинах 22-27 м фитобентос почти полностью исчезает, что связано с доминированием песчаных и илистых грунтов, ракушечника, а также с ослаблением фотосинтетически активной радиации до значений, близким к компенсационной точке фотосинтеза. ОПП на такой глубине редко больше 1%. Там, где позволяет прозрачность воды и характер грунта, фитоценозы развиваются и достигают биомассы 0,5-2 кг/м2. Доминируют Сodium vermilara (300-350 г/м2) или Phyllophora nervosa (200-800 г/м2). Второй – четвертый ярус развиты слабо и представлены теми же видами, что и на предыдущей глубине.
В Северо-восточном районе, в районе полуострова Абрау изобаты проникновения растительности различны, причем на антропогенно нарушенных участках, в связи с меньшей прозрачностью воды и заилением грунта, макрофитобентос исчезает на меньших глубинах. Так, в Новорссийской бухте исчезновение растительности происходит на глубинах до 15 м. В чистых экотопах глубина проникновения растительности увеличивается в 2 раза (до 32 м у м. Б. Утриш). Растительность участка г. Туапсе – г. Геленджик исчезает на глубине около 9,5 м, в связи с изменением грунта с каменистого и скально-валунного на песчаный или песчано-илистый (Афанасьев, 2005).
Фитобентос российского сектора Юго-восточного гидроботанического района Черного моря, в отличие от Северо-восточного, представлен относительно небольшим числом в основном короткоцикличных тепловодных и эврибионтных видов отделов зеленых и красных водорослей, встречающихся по всей акватории Черного моря. В целом по району на глубине до 1 метра в полосе прибрежной гальки, растительность отсутствует. На глубине 2-3 м основу донных растительных сообществ составляют развивающиеся на твердом грунте ассоциации Cystoseiretum ulvosum и Cystoseiretum dilophoso-cladostephosum, аналогичные приведенным выше, с ОПП от 30 до 85% и биомассой 1,6 кг/м2. Опреснение и повышенная сапробность воды обусловливает преобладание во втором ярусе мезо - и полисапробных зеленых и красных водорослей родов Enteromorpha, Chaetomorpha, Cladophora, а также Ulva rigida, Callithamnion corymbosum, Ceramium rubrum, ПП которых может быть весьма значительным. В качестве ассектаторов в сообществе могут присутствовать Stilophora rhizodes, Dilophus fasciola. Эпифитируют с незначительной биомассой в основном Polysiphonia subulifera и Laurencia obtusa. К изобате 5 м средняя биомасса водорослей снижается в среднем до 1,1 кг/м2. На исследованной акватории растительность исчезает в среднем на глубине 6 м (Афанасьев, 2009). Слабое развитие фитобентоса южнее г. Туапсе определяется в первую очередь большим количеством рек, впадающих здесь в море, выбросом в море значительного количества аллювия, существенно снижающего прозрачность опресненных прибрежных вод и создающего предпосылки для доминирования галечного и илисто-песчаного грунтов (Пешков, 2000; Петров, 2001; Афанасьев, 2009).
Сравнение современной структуры растительности, с таковой, описанной -Гутник (1964, 1970, 1975) по результатам работ г. и (1998) в 70-80-х гг. ХХ в. показывает, что к 80-м гг. произошло исчезновение в Северо-восточном и Юго-восточном гидроботанических районах зарослей зостеры, снижение биомассы основных видов и частоты встречаемости олигосапробных видов водорослей. Так, сравнивая наши данные с данными (1963) и (1964), обнаруживаем, что зостера ранее доминировала на песке в Анапской бухте на глубинах от 0,6 до 2,2 (Zostera noltii) и от 3 до 17 м (Z. marina) со средней биомассой 380 г/м2 и максимальной – 800 г/м2. В настоящее время, здесь редко встречаются отдельные угнетенные куртины Z. noltii с максимальной биомассой до 85-110 г/м2. В Новороссийской бухте зостера встречалась в виде отдельных пятен в северо-восточной части порта, в Галацкой бухточке и в районе Кабардинки со средней биомассой 150 г/м2 (Калугина, 1964). Сейчас лишь отдельные экземпляры зостеры можно встретить в районе Суджукской косы. Если на участке г. Туапсе – г. Сочи на глубине более 3-5 м песчаная сублитораль ранее была покрыта изреженными лугами Zostera noltii, то теперь совсем лишена растительности. Общая биомасса обоих видов зостер достигала нескольких десятков тысяч тонн сырой массы. Причиной элиминации зарослей зостеры могла послужить как эпифитотия, вызванная лабиринтуломицетами, как это произошло с Zostera marina в 30-х гг. ХХ в. (Морозова-Водяницкая, 1936), так и возрастающие эвтрофикация и загрязнение прибрежной зоны моря (Громов, Афанасьев, 2002).
За 50 лет средняя по побережью биомасса цистозиры на глубине 1 м уменьшилась в среднем в 3 раза, на глубине 3 м – в 2,6 раза, на глубине 5 м – в 2,8 раза, на глубине 10 м – в 3,3 раза; средняя длина таллома цистозиры также уменьшилась в среднем в 2-2,5 раза. Густые заросли цистозир у открытого берега Северо-восточного гидроботанического района ранее располагались до глубины 20 м (Калугина, 1964), ныне – до глубины 10-12 м. Еще более значительно уменьшилась биомасса филлофоры: если для участка от м. Идокопас до м. Тонкого она составляла в гг. на глубинах 10-20 м в среднем 773 г/м2 (Калугина, 1964), то в гг. – всего 58 г/м2. Уменьшение встречаемости преимуществено олигосапробных видов водорослей, таких как Dictyota dichotoma, Dilophus fasciola, Lophosiphonia reptabunda, L. obscura, Apoglossum ruscifolium, Acrochaetium secundatum, Grateloupia dichotoma, Gracilaria gracilis, Lomentaria clavellosa и ряда других особенно значительно в Юго-восточном гидроботаническом районе. Отмечено, что биомасса доминанта второго яруса ассоциации Cystoseiretum dilophoso-cladostephosum – Cladostephus spongiosus f. verticillatus – в районе снизилась сг/м2 в 1993 г. до 1-13 г/м2 в 2006 г.
Обращает на себя внимание факт почти полного исчезновения в Юго-восточном районе уже к 90-м гг. ХХ в. некоторых видов водорослей (Phyllophora crispa, Apoglossum ruscifolium, Chondria capillaris, C. dasyphylla и др.), что привело к флористическому обеднению ассоциаций. Так, на некоторых разрезах отмечаются практически чистые заросли цистозиры бородатой с минимумом (2-3) сопутствующих видов.
Таким образом, фитобентос Северо-восточного и Юго-восточного районов претерпел существенные изменения структуры, связанные с обеднением флористического состава сообществ, исчезновением ассоциаций зостеры, значительным (в 2-10 раз) снижением биомассы других ценообразующих видов, снижением глубины проникновения растительности.
Растительность Таманского полуострова и Керченского пролива
У м. Тузла, Панагия и Железный Рог у уреза воды песчаные грунты лишены растительности. Лишь к 0,3 м глубины появляется Zostera noltii, которая редкими куртинами простирается до глубины 5 м. Биомасса куртин не превышает 800 г/м2, при ПП всего в 10-20%. Единично на выступающих поверхностях камней с микрорельефом до 1,5 м встречаются преимущественно Callithamnion corymbosum, Enteromorpha clathrata, Ceramium rubrum и Polysiphonia opaca.
Вся центральная часть Таманского залива с глубинами 3,5-6,0 м занята двухъярусной ассоциацией морских трав Zosteretum marinae potamagetosum. На грунте эта ассоциация выглядит в виде чередующихся полос и пятен из морской травы Zostera marina и рдеста Potamogeton pectinatus с ОПП 25-100%, причем ПП зостеры почти в 3 раза превышает покрытие рдеста. Фитомасса ассоциаций колеблется от 280 до 5140 г/м2. По мере уменьшения глубины в поясе от 3,5 до 0,5 м развивается двухъярусная ассоциация Zosteretum subpurum, верхний ярус которой высотой 70 см образует Z. marina, нижний – Z. noltii. ОПП этой ассоциации колеблется от 20% до 90% при фитомассе г/м2. Самая прибрежная часть Таманского залива с плотным песчаным грунтом занята низкорослой одноярусной ассоциацией Zosteretum noltii purum c фитомассой до 840 г/м2. Из водорослей в Таманском заливе наиболее обычны Cladophora laetevirens со значительной биомассой (до 400 г/м2), Chaetomorpha crassa, Enteromorpha clathrata, Ceramium rubrum (с биомассой до 200 г/м2), Chondria capillaris (= C. tenuissima), эндемик Enteromorpha maeotica. Вдоль всего восточного берега косы Чушки и у западного берега залива в зоне псевдолиторали на плотном песчаном грунте развивается ассоциация тростника Phragmitetum purum с ОПП до 100%. Средняя фитомасса этой ассоциации достигает 4100 г/м2 при плотности тростника до 60 экз/м2 (Громов, Шевченко, Афанасьев, 2002).
Водная растительность прибрежной акватории юго-восточной части Азовского моря представлена фрагментарными сообществами, преимущественно высших растений. Глубже изобаты 6,5 м, в зоне распространения илов, макрофитобентос не встречается (Громов, 1999), что подтверждается и нашими исследованиями (Афанасьев, 2006). Наибольшее видовое разнообразие фитобентоса обнаружено у Керченского предпроливья. Основной тип растительности здесь – Thallassophycion – сообщества морских водорослей, класс формаций Thallassophycion malacochthonophyceae – сообщества свободноплавающих и прикрепленных морских водорослей на мягком естественном грунте, доминирует ассоциация Ceramietum subpurum, относящаяся к группе формаций Rhodophyta – красные водоросли. В собственно море доминирует тип Thallassopoion – сообщества морских трав, класс формаций Thallassopoion malacochthonophyceae – сообщества морских трав на мягком естественном грунте, группа формаций Thallasopoion magnoliophyceae, представленная ведущими ассоциациями Zosteretum subpurum и Zosteretum noltii purum.
В лиманах на илистом и илисто-песчаном грунте доминирует высшая водная растительность, представленная в основном ассоциациями Phragmitetum subpurum, Potametum purum, относящимися к группе формаций Magnoliophyceae, класса формаций Limnophycion malacochthonophyceae, типа сообществ Limnopoion. Тростнику сопутствуют около 30 таксонов макрофитов пресноводного, пресноводно-солоноватоводного и солоноватоводного комплексов. На открытых участках на илистых и илисто-песчаных грунтах преобладают моно - и олиговидовые ассоциации Ceratophyllum demersum, Potamogeton pectinatus, реже P. crispus, P. perfoliatus, Vallisneria spiralis, Miryophyllum spicatum. Общими для всех лиманов является тростник и уруть. В пресных лиманах к ним прибавляется рогоз, роголистник и рдесты, в соленых – заникелия, зостера, многочисленные виды зеленых водорослей родов Enteromorpha, Chaetomorpha, Cladophora. Динамика биомассы за последние 30 лет, приведенная в работе (2006) свидетельствует о постепенном зарастании лиманов и плавен и сокращении чистого зеркала большинства водоемов. Так, общая сырая фитомасса погруженной высшей водной растительности в Кубанских лиманах по данным ныне оценивается в 2,043 млн. тонн, тогда как в гг. оценивалась в 1,193 млн. тонн (Шехов, 1971; Цуникова, 2006). Таким образом, биомасса водной растительности в целом по Кубанским лиманам увеличилась в 1,7 раза, что связано с ухудшением мелиорации лиманов.
При сопоставлении результатов наших исследований с данными (1971), отмечено практически полное исчезновение харово-урутьево-гребенчатордестовых и телорезово-кувшинковых сообществ лиманов. Роголистниково-харовые сообщества заменились на чистые роголистниковые сообщества либо сообщества роголистника с примесью валлиснерии. Кроме того, отмечается существенное снижение видового разнообразия фитобентоса лиманов. Так, если (1971) отмечает в лиманно-плавневой зоне 55 видов высших водных растений и 20 видов водорослей, то нами отмечено лишь 28 видов высших растений и 18 видов водорослей.
В силу сокращения усилий по мелиорации лиманов, в последние годы возросло количество водоемов, находящихся на конечных стадиях сукцессий (преимущественно, рогозово-тростниковые сообщества). Таким образом, облик современного фитобентоса лиманно-плавневой зоны определяется, во-первых, заболачиванием и зарастанием лиманов преимущественно сообществами рдестов, рогоза и тростника, что обуславливается сокращением объема проводимых мелиоративных мероприятий, и, во-вторых, обеднением видового состава за счет выпадения олигосапробных видов при загрязнении и эвтрофикации водоемов.
Анализ многолетней динамики и проблемы классификации растительности Азово-Черноморского шельфа
В разделе приводятся классификационные схемы растительности Азовского и Черного морей, предложенные разными авторами (Морозова-Водяницкая, 1959; Петров, 1961; Калугина-Гутник, 1975; Громов, 1998, 2000). Отмечаются положительные и отрицательные стороны различных классификационных построений. Наше следование классификации -Гутник (1975) с дополнениями (1998, 2000) определяется удобством сравнения данных по современному состоянию растительного покрова Черного моря с данными, полученными ею в 60-х гг. ХХ века и отражающими нативное состояние растительности до начала серьезных антропогенных преобразований.
На текущий момент в прибрежных экосистемах российского шельфа Черного моря насчитывается 27, Азовского моря – 33 растительные ассоциации. Современный анализ распространения ассоциаций позволяет выделить те, которые значительно сократили площадь распространения по всему российскому шельфу. Это ассоциации Feldmannietum purum, Chondrietum laurenciosum, Polysiphonietum subpurum, Antithamnietum subpurum (ранее относившиеся к группе региональных), Gracilarietum neriosum, Zosteretum noltii purum, Zosteretum marine-polysiphoniosum, Lophosiphonietum subpurum, Ruppietum marinae purum, Potametum pectinati purum (ранее относившиеся к группе локальных). Все указанные ассоциации являются олигосапробными. Еще одна олигосапробная ассоциация – харовых водорослей Lamiprothamnium papulosum и Chara vulgaris (Lamiprothamnietum subpurum), описанная (1935) и -Гутник (1975) для Суджукской лагуны Новороссийской бухты – по нашему мнению, вообще более не встречается.
Анализируя вертикальное распределение ассоциаций, необходимо указать на то, что если по данным -Гутник (1975) нижняя граница фитобентоса Черного моря проходила на глубине 80-90 м (а согласно (1908) – на глубине 125 м), то в настоящее время на северо-кавказском шельфе глубже 32 м водоросли-макрофиты не встречаются. Причиной является, очевидно, снижение прозрачности воды во всем Черном море, произошедшее в последние два-три десятилетия. В связи с уменьшением глубины проникновения водорослей произошло сокращение ширины зарослей фитобентоса, а нижние границы каждого этажа сублиторальной зоны переместились в сторону меньших глубин. Так, на участке с максимальной глубиной проникновения донной растительности (полуостров Абрау) нижняя граница Iго этажа сублиторальной зоны сместилась с изобаты 5 м до изобаты 4 м, IIго этажа – с глубин 5-15 м до глубин 4-10 м, IIIго этажа – с глубин 15м до глубин 10м, IVго этажа (по -Гутник (1975) – I этаж сциафильной растительности) – с глубинм до глубинм, последнего этажа – с глубин 60-90 м до глубин 30м.
Снижение прозрачности воды привело также почти к полному исчезновению самых глубоководных ассоциаций (Chondrietum laurenciosum, Gracilarietum neriosum, Polysiphonietum subpurum, Antithamnietum subpurum) и сужению ширины ассоциаций средних этажей (Cystoseiretum dilophoso-cladostephosum, Cystoseiretum phyllophorosum, Phyllophoretum purum).
Все ассоциации, ставшие более редкими по сравнению с 60-ми гг. ХХ в., относятся к группе олигосапробных. Данное обстоятельство выдвигает эвтрофикацию шельфовых экосистем Черного моря (вместе со снижением прозрачности воды и заилением дна) на одно из первых мест по влиянию на структуру и состав макрофитобентоса.
Следует особо отметить тот факт, что если на мелководье олигосапробные фитоценозы замещаются мезо - и полисапробными, то при деградации глубоководных ассоциаций дно моря обнажается – на северо-кавказском шельфе Черного моря нет ни одной β-мезосапробной и полисапробной ассоциации, которая развивалась бы глубже I этажа сублиторальной зоны (табл. 3).
Таким образом, наиболее уязвимыми на российском шельфе Черного моря являются глубоководные фитоценозы и фитоценозы аккумулятивных ландшафтов, занятых сообществами морских трав и харовых водорослей. Сообщества первых двух этажей сублиторали на твердых грунтах, более разнообразные по структуре и занимающие разные экологические ниши, в целом и более устойчивы к антропогенному воздействию. Наиболее широкой экологической валентностью и продуктивностью обладают асс. формации Cystoseireta, имеющие достаточно мощный адаптационный потенциал, позволяющий им занимать акватории на глубинах от 0,1 до 10-15 м, различающихся степенью прибойности и сапробности.
Таблица 3. Экологическая характеристика морских ассоциаций класса формаций сообществ твердых грунтов*
сапробность единицы вертикального деления | олигосапробные ассоциации | мезосапробные ассоциации | полисапробные ассоциации | |
супралиторальная зона (до 1 м выше уреза воды) | Feldmannietum purum** | Bangietum purum | Urosporetum purum | |
псевдолиторальная зона (0-0,1 м) | Scytosiphonietum purum | Porphyretum purum | ||
сублиторальная зона | I этаж (0,1-4 м) | Dilophetum ceramioso – polysiponiosum, Cystoseiretum dilophoso – cladostephosum, Cystoseiretum dilophosum | Cystoseiretum ulvosum, Ulvetum ceramiosum, Cladophoretum enteromorphosum, Enteromorphetum linza polysiphoniosum opaca | Enteromorphetum intestinalis ceramiosum rubrum, Enteromorphetum intestinalis purum |
II этаж (4-10 м) | Cystoseiretum dilophoso – cladostephosum, Cystoseiretum phyllophorosum, Chondrietum laurenciosum, Gracilarietum neriosum | Cystoseiretum phyllophorosum | - | |
III этаж (10-25 м) | Codietum phyllophorosum, Phyllophoretum purum, Chondrietum laurenciosum, Polysiphonietum subpurum | Codietum phyllophorosum | - | |
IV этаж (25-30 м) | Polysiphonietum subpurum | - | - | |
V этаж (30-35 м) | Antithamnietum subpurum | - | - |
* - ассоциации в каждой ячейке даны в порядке приуроченности к участкам с менее интенсивным движением воды (снижение прибойности), каждый последующий этаж также характеризуется меньшей интенсивностью движения воды
** - жирным выделены ассоциации, ставшие сегодня более редкими, по сравнению с 60-ми годами ХХ века
В разделе приводятся данные по ценотической характеристике ассоциаций и их продуктивности.
Флора макрофитов Азово-Черноморского шельфа и ее изменение под влиянием антропогенного фактора
На основании результатов собственных исследований, дополненных анализом литературных данных, составлен современный список макрофитов российского сектора Азовского и Черного морей и определена их встречаемость. Учитывая сине-зеленые и желто-зеленые водоросли, образующие видимые невооруженным глазом скопления, всего в Азово-Черноморской флоре в указанных границах описано 313 видов макрофитов и 321 внутривидовой таксон, относящихся к 8 отделам: Cyanophyta (27 видов), Chlorophyta (65 видов), Charophyta (13 видов), Xanthophyta (1 вид), Rhodophyta (100 видов), Phaeophyta (47 видов), Polypodiophyta (2 вида) и Magnoliophyta (57 видов). Анализ динамики встречаемости водорослей-макрофитов в Черном море с 1975 года по настоящее время, показывает, что в целом в российском секторе Черного моря 6% видов стали встречаться чаще (преимущественно зеленые водоросли), стали более редкими 23% видов, исчезло 5% видов (в основном бурые водоросли). Все исчезнувшие виды являлись редкими. Из зеленых водорослей это Caulerpa prolifera (Forsk.) Lamour.; из бурых – Myrionema balticum (Reinke) Foslie, Microspongium gelatinosum Reinke, Pseudolithoderma extentum (Crouan) S. Lund., Streblonema sphaericum Derb. et Sol., Myriactula arabica (Kutz.) Feldm., Acinetospora crinita (Carm.) Kornm., Punctaria plantaguinea (Roth) Grev., Asperococcus bullosus Lamour.; из красных – Kylinia polyblasta (Rosenv.) Papenf.
В диссертации приводится флористический анализ современного списка видов. Показано, что сукцессии макрофитобентоса российской части Азово-Черноморского шельфа включают в себя изменение структуры флоры макрофитов. Наиболее существенные изменения флоры за последние 40 лет произошли на шельфе Черного моря и заключались не только в изменении встречаемости видов в сторону преобладания короткоцикличных, тепловодных, эврибионтных видов, но и в полном выпадении из флоры 10 видов макрофитов. Данное обстоятельство существенно отличает российский берег Черного моря от турецкого, румынского и болгарского. Если там происходит увеличение числа таксонов в результате продолжающейся медитерранизации флоры и проникновения новых видов (Мильчакова, 2002, 2003, 2004; Заика и др., 2004; Milchakova et al., 2005), то для российского шельфа в настоящее время характерен тренд уменьшения видового богатства макрофитов.
Нами предлагается выделить для Азово-Черноморского побережья, на основе морфо-физиологических особенностей донного растительного покрова 4 типа флор (Громов, Афанасьев, 2002): 1) естественные флоры, складывавшиеся в чистых экотопах, в отсутствие антропогенного пресса (данный тип флор в современной период, скорее всего, утрачен); 2) полуестественные обогащенные флоры, формирующиеся в местах незначительной рекреационной нагрузки, в акваториях возле небольших населенных пунктов; включают олиго - и α-мезосапробные виды водорослей; 3) урбанофлоры, для которых целесообразно выделить два подтипа: а) сапробные флоры, включающие мезо - и полисапробные виды, характерные для участков со значительными рекреационными нагрузками; отражают, в первую очередь, высокую степень сапробности и б) собственно урбанофлоры, складывающиеся, как правило, в портах, характеризующихся загрязнением токсическими веществами (тяжелыми металлами, нефтяными углеводородами и т. п.), а так же низкой прозрачностью вод, заилением грунта. Подобные флоры крайне обеднены. Особо необходимо выделить 4) тип – техногенные флоры, развивающиеся в экотопах, сформированных в процессе хозяйственной деятельности и не имеющих природных аналогов; перифитон (флора пирсов, буев, молов, днищ кораблей и т. д.). Репрезентативны по отношению к флоре региона полуестественные обогащенные флоры; каждый из типов трансформированных флор имеет обедненный видовой состав (кроме второго типа) и ни один из них не охватывает все виды флоры региона полностью; для последних трех типов флор, характерны явления, присущие синантропизации растительного покрова: унификация, замена эндемиков – космополитами, стенотопных видов – эвритопными, упрощение структуры сообществ.
Глава 4. Современное состояние популяций доминирующих видов водорослей Черного моря
При изучении возрастной структуры популяций цистозиры кустистой и бородатой обнаружена приуроченность растений разных возрастов к разным глубинам. На мелководье популяции представлены молодыми растениями, с увеличением глубины увеличивается доля старых слоевищ. Так, на достаточно чистых, открытых участках побережья (Абрауский полуостров) на глубине 1 м средний возраст растений составляет 2-4 года, а на глубине 3 м – 4-7 лет, и далее, с ростом глубины практически не меняется (рис. 2, 3). В защищенных районах, без подтока сточных вод, возраст цистозиры максимален: на глубине 1 м основная часть популяции представлена 5-6-летними растениями, глубже – 6-10-летними (Афанасьев, Степаньян, 2001).
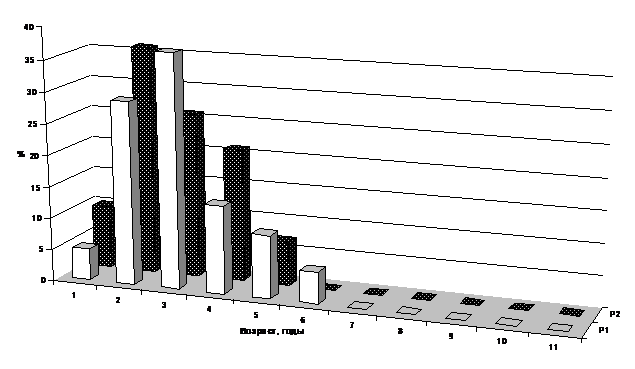
Рис. 2. Возрастная структура популяций цистозиры бородатой на глубине 1 м (Р1 – контрольный участок (СОЛ "Лиманчик", Абрауский полуостров), Р2 – эвтрофицированный участок (м. Любви, Новороссийская бухта)
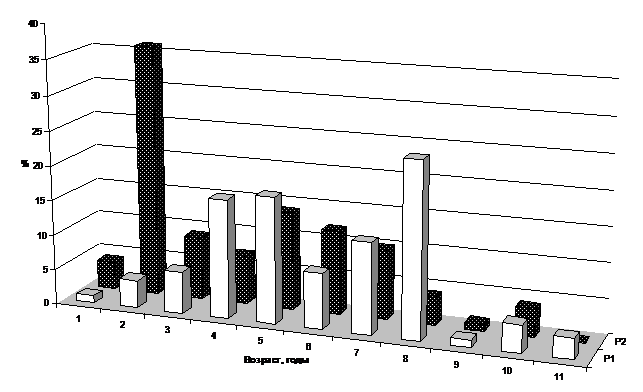 Рис. 3. Возрастная структура популяций цистозиры бородатой на глубине 3 м (Р1 – контрольный участок (СОЛ "Лиманчик", Абрауский полуостров), Р2 – эвтрофицированный участок (м. Любви, Новороссийская бухта)
Рис. 3. Возрастная структура популяций цистозиры бородатой на глубине 3 м (Р1 – контрольный участок (СОЛ "Лиманчик", Абрауский полуостров), Р2 – эвтрофицированный участок (м. Любви, Новороссийская бухта)
Для районов с выпуском бытовых стоков и интенсивным волнением характерно снижение среднего возраста растений по всем глубинам. Так, у м. Любви на глубине 1м нет растений старше 5-летнего возраста, а в основном доминируют 2-3-летние особи; на 3-х м средний возраст популяции повышается до 4-х лет (рис. 2, 3).
Наши данные совпадают с выявленными ранее закономерностями распределения возрастных групп цистозиры по глубинам (Ковардаков и др., 1985).
Подробное морфологическое и физиолого-биохимическое исследование 3 ценопопуляций Cystoseira crinita, обитающей на глубине 0,2 м, в экотопах, отличающихся уровнем эвтрофикации (Новороссийская бухта (м. Шесхарис, м. Любви) и Абрауский полуостров (м. Большой Утриш) позволило выявить, что в условиях эвтрофикации средняя масса водорослей (как и прямо пропорциональная ей площадь поверхности) в 3 раза выше (притом, что достоверных различий в возрасте у данной выборки с популяциями других экотопов нет). Поскольку биомасса водорослей растет быстрее, чем площадь поверхности, удельная площадь поверхности (отношение площади поверхности к биомассе) снижается. Увеличение биомассы всего таллома в условиях эвтрофикации осуществляется преимущественно за счет интенсификации развития адвентивного комплекса (увеличения количества (более чем в 3 раза) и массы (более чем в 20 раз) адвентивных ветвей). Снижение удельной площади поверхности цистозиры кустистой в условиях эвтрофикации и загрязнения приводит к уменьшению физико-химического контакта растения со средой, из чего следует, что перераспределение интенсивности развития структур водоросли в таких условиях – часть адаптационной стратегии. В отличие от комплекса адвентивных ветвей, количество и масса основных (верхушечных) ветвей – более стабильный признак, что указывает на то, что основной комплекс не принимает существенного участия в адаптации водоросли (масса основных ветвей практически одинакова у растений разных экотопов). Таким образом, отношение масс адвентивных и основных ветвей служит хорошим критерием для оценки степени эвтрофности водоема и отражает усиление роли адвентивного комплекса цистозиры в условиях эвтрофикации (Афанасьев, 2008).
Обнаружено, что в условиях и эвтрофикации, и нефтяного загрязнения интенсивность хемилюминесценции, пероксидазной активности, содержание ТБК-активных продуктов, каротина и ксантофиллов в молодых основных ветвях 1-го порядка цистозиры достоверно меняется, что указывает на изменение окислительно-восстановительного статуса водоросли. Кроме того, в таких условиях наблюдается снижение концентрации хлорофилла “а” и увеличение содержание хлорофилла “с”. Однако, изменение концентрации зеленых пигментов в основных ветвях не достаточно скомпенсировано: уменьшению содержания хлорофилла “а” на 8,9 мг/100 г, соответствует увеличение концентрации хлорофилла “с” лишь на 1,6 мг/100 г и менее. Таким образом, вклад каждой отдельной ветви в общий фотосинтез слоевища снижается. Адаптивный смысл этого явления становится понятен, если учесть, что снижение концентрации зеленых пигментов, возможно, способствует снижению риска индукции переокисления липидов. В таком случае, снижение интенсивности фотосинтеза каждой отдельной ветви, оцениваемой по общему содержанию хлорофиллов, на уровне целого организма компенсируется увеличением массы и количества ветвей.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
