
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
При сопоставлении полученных нами данных с результатами исследований, проведенных в 60-х годах, обнаружено, что произошло существенное уменьшение площади произрастания и общей биомассы ценообразующих видов – зостеры, цистозиры и филлофоры (рис. 4).
Из рисунка 4 следует, что снижение общей биомассы цистозиры до начала 80-х годов прошлого столетия слабо выражено. Это было обусловлено локальным характером замены цистозировых ценопопуляций в местах, подвергающихся интенсивной эвтрофикации и загрязнению (Анапа, Новороссийск, Геленджик, Туапсе), ассоциациями эфемерных видов родов Enteromorpha, Ulva, Chaetomorpha, Spongomorpha, Cladophora, Ectocarpus, Ceramium, Polysiphonia и некоторых других.
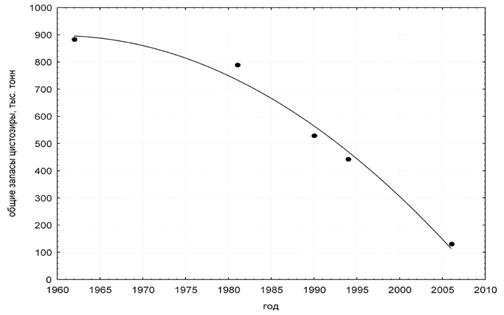
Рис. 4. Общая биомасса цистозиры на российском побережье Черного моря с 60-х гг. XX века по настоящее время (по данным -Гутник (1964, 1975, 1994); (1991); (1998); собственным данным)
С конца 80-х гг. темпы снижения общей биомассы цистозиры ускорились, что было обусловлено поднятием нижней границы ее зарослей и сужением их ширины на российском шельфе в целом.
Причиной явления считается снижение в полтора-два раза прозрачности вод Черного моря, наблюдаемое с конца 70-х годов (Техногенное…, 1996). Результаты проведенного нами корреляционного анализа указывают на существование связи между прозрачностью воды на российском шельфе Черного моря и общей биомассой цистозиры: коэффициент корреляции составляет 0,92 (при р < 0,05). Таким образом, одной из основных причин сокращения зарослей цистозиры и снижения глубины распространения растительности на российском шельфе Черного моря действительно является снижение прозрачности воды. Помимо увеличения содержания органической и минеральной взвеси, эвтрофикации и интенсивного развития планктонных организмов (Калугина-Гутник, 1994; Маньковский и др., 1996; Кукушкин и др., 2004, 2006). в качестве причины снижения прозрачности воды называется вселение в 1982 г. в Черное море гребневика Mnemiopsis leidyi A. Agassiz (Кучерук и др., 2002; Максимова, Лучина, 2002; Maximova, Moruchkova, 2005). Кроме того, увеличение содержания в воде взвешенного вещества могло произойти, в том числе, из-за нарушения естественного хода седиментогенеза на шельфе, вызванного резким снижением численности организмов, осуществляющих седиментацию взвеси - моллюсков-фильтраторов, и в первую очередь устриц и мидий. Запасы мидий в советском секторе Черного моря по некоторым оценкам сократились с 64,5 млн. т. в 1933 г. до 3,5-9,6 млн. т. в 60-70-е гг. (Кучерук и др., 2002; Елецкий, 2006). В районе Кавказа основной фактор снижения численности устриц и мидий – выедание хищным брюхоногим моллюском - вселенцем Rapana thomassiana Gross (Фроленко и др., 2005; Елецкий, 2006). Полное исчезновение устриц и существенное снижение запасов мидии после вселения рапаны вполне могло стать причиной уменьшения самоочистительной способности прибрежной зоны моря и повышения доли взвешенного органического и минерального вещества, что привело, в конечном счете, к снижению прозрачности воды у берега и сужению зарослей макрофитов. Указанная причина сужения зарослей повлекла за собой и сокращение общей биомассы других видов, в частности филлофоры – ее биомасса оценена в 15,8 тыс. тонн, при оценке -Гутник (1964) в 31 тыс. тонн.
Итак, за последние 50 лет на российском шельфе Черного моря произошли существенные изменения общей биомассы макрофитобентоса, обусловленные снижением прозрачности воды, эвтрофикацией и загрязнением шельфовой зоны, инвазиями чужеродных видов (гребневик Mnemiopsis leidyi, брюхоногий моллюск Rapana thomasiana). В целом, почти полностью исчезли ассоциации зостеры, общая биомасса цистозиры снизилась более чем в 6 раз, филлофоры – в 2 раза, а ульвы – несколько увеличилась. Если в 50-х, 60-х гг. ведущими видами – ценообразователями в альгофлоре всего Черного моря являлись филлофора, цистозира и зостера (Генералова, 1952; Тренина, 1959; Морозова-Водяницкая, 1927, 1959; Киреева, 1964; Калугина и др., 1967), то теперь – цистозира и лишь локально филлофора (Афанасьев, 2008, 2009).
Оценка изменения экологической роли макрофитов в прибрежье Черного моря возможно на основании сопоставления общей продукции живого вещества на шельфе до начала интенсивных антропогенных преобразований и в последнее время (согласно Зенковичу, 1963; Финенко, 1967, 1979; Грезе, Федориной, 1979; Сорокину, 1982; Зайцеву, 1998, 2000; Елецкому, 2006; Отчетам АзНИИРХ…, 2005, 2007, 2008; собственным данным). Из результатов проведенного анализа следует, что прибрежные экосистемы Черного моря претерпевают существенные перестройки под влиянием антропогенного фактора – общая продукция фито - и зоопланктона в последние два десятилетия увеличилась, что является следствием в первую очередь значительной эвтрофикации прибрежной зоны моря, продуктивность донных биоценозов – снизилась. Так, если ранее общая продукция фитобентоса в пределах прибрежной зоны до глубины 100 м на восточном побережье Черного моря была всего в 2.5 раза ниже, чем общая продукция фитопланктона, то по данным на сегодняшний день – почти в 20 раз меньше. Таким образом, основной поток вещества и энергии в прибрежных экосистемах в настоящее время направлен через планктонные биоценозы. По данным (2007), аналогичные процессы увеличения первичной продукции за счет усиления развития фитопланктона характерны и для Азовского моря.
Наиболее экологически обоснованным способом восстановления зарослей макрофитов является внесение в фотический слой воды искусственных субстратов, обрастающих водорослями. Так, в Юго-восточном гидроботаническом районе Черного моря в условиях доминирования подвижных грунтов – песка, гальки, ила – роль перифитона в псевдолиторали весьма существенна. При ОПП 35-90%, сообщества водорослей перифитона достигают средней биомассы 900 г/м2 (максимум 1625 г/м2), что выше, чем на естественном грунте. Для растительности искусственных субстратов черноморского побережья существует ряд специфических закономерностей, установленных нами в результате исследований: во-первых, слабое развитие на искусственных субстратах корковых водорослей; во-вторых, интенсивное развитие видов, образующих дерновины (Corallina officinalis, C. elongata, Gelidium crinale, G. spinosum, Spongomorpha arcta); в-третьих, редкая встречаемость некоторых обычных на естественных субстратах видов водорослей (Dilophus fasciola, Padina pavonia, некоторые виды родов Polysiphonia и Laurencia). Всего флора перифитона представлена на российском шельфе Черного моря 80 видами водорослей-макрофитов. В Юго-восточном районе часть видов водорослей встречается только в составе обрастаний, что определяется регулярной отсыпкой песка на пляжи и сокращением площадей экотопов, пригодных для макрофитов. Это еще раз подтверждает, что одним из основных средств сохранения биоразнообразия (если не единственным) является расстановка вдоль побережья искусственных субстратов.
Перифитон юго-восточного района Азовского моря беден в систематическом отношении и представлен только 10 таксонами водорослей-макрофитов. На глубинах до 0,8 м в Темрюкском заливе Азовского моря на искусственных субстратах развиваются монодоминантные чистые сообщества водорослей, с максимальной весенней биомассой от 140 до 230 г/м2. В обрастании субстратов доминируют два вида зеленых водорослей: Enteromorpha prolifera и E. linza, что обусловливает невысокие значения индекса биоразнообразия. Отсутствие, за редкими исключениями, крупных, многолетних видов водорослей, не позволяет формировать многоярусные, полидоминантные ассоциации. Напротив, интенсивное развитие типичных r-стратегов – мелкоразмерных, сезонных видов водорослей-макрофитов (относящихся в основном к родам Enteromorpha, Cladophora и Ceramium) – приводит к наблюдаемому релаксационно-колебательному характеру изменений плотности их ценопопуляций. Наибольшая функциональная активность перифитона приходиться на конец весны – начало лета, когда наблюдаются максимальные значения биомасс водорослей-макрофитов, в это же время целесообразно внесение в фотический слой дополнительных поверхностей, обрастающих макрофитами, причем на глубину не более 1 м.
В процессе сукцессионных изменения сообществ обрастания твердых субстратов в Азовском море есть ряд определенных закономерностей. В частности, происходит смена преобладания в видовом составе и биомассе фитобентоса на зообентос. Согласно нашим наблюдениям, сообщества искусственных субстратов в Азовском море проходят следующие последовательные стадии: фито-балянусная – балянусная – балянусно-малако-мшанковая – малако-мшанковая и сложноколониальная. Последняя стадия является климаксной, характеризуется наибольшим видовым разнообразием и устойчивостью. Такие сообщества формируются на субстратах обычно не ранее, чем через 1-3 года.
Таким образом, восстановление популяций основных в экологическом и хозяйственном отношении видов (Cystoseira barbata, C. crinita, Codium vermilara, Ulva rigida, Phyllophora crispa) на Азово-Черноморском шельфе России с помощью гидробиотехнических конструкций возможно, так как все указанные виды входят во флору искусственных экотопов и формируют здесь зрелые популяции.
На основании проведенной оценки продуктивности и общей биомассы макрофитобентоса побережья Азовского и Черного морей, разработаны рекомендации по сохранению, восстановлению и рациональному использованию макрофитов.
Заключение
Среди компонентов морских экосистем, подвергшихся в результате антропогенных действий глубоким перестройкам, одним из наименее изучаемых и изученных объектов является макрофитобентос. Для Черного моря, анализ имеющихся данных делает возможным выделение нескольких периодов преобразования фитобентоса за последнее столетие.
Первый период, наиболее длительный по времени, связан с началом интенсивной антропогенной деятельности на побережье. Он характеризовался активным прибрежным строительством, началом эвтрофикации прибрежных акваторий, формированием локальных мезо - и полисапробных ассоциаций (20-е – конец 60-х гг. ХХ в.).
В 70-е – 80-е гг. начинается второй период – интенсивной эвтрофикации акваторий, сопряженной с началом массированного загрязнения прибрежных вод нефтепродуктами, тяжелыми металлами, сельскохозяйственными ядохимикатами. Все это повлекло за собой локальное изреживание зарослей макрофитов, частичное исчезновение фитобентоса на некоторых глубинах (например, у нефтебазы
Шесхарис в конце 60-х – начале 70-х гг.), продолжение распространения мезо - и полисапробных ассоциаций на шельфе.
Третий (современный) период, начавшийся в 90-е гг. ХХ в., характеризуется наибольшей напряженностью. Он связан в первую очередь со значительным снижением прозрачности морской воды, мощной эвтрофикацией и загрязнением всего шельфа, изменением характера донных отложений. Современные изменения включают почти 3-х кратное сужение полосы водной донной растительности на северо-кавказском шельфе Черного моря, 2-6-кратное уменьшение биомассы ведущих видов водорослей, практически полную деградацию олигосапробных и глубоководных ассоциаций, существенное снижение биоразнообразия донных сообществ (в том числе исчезновение некоторых видов).
В целом, на Азово-Черноморском шельфе причинами антропогенных сукцессий макрофитобентоса являются загрязнение и эвтрофикация, снижение прозрачности воды и изменение характера грунта. При этом, для Юго-восточного гидроботанического района Черного моря основными причинами указанных сукцессий являются эвтрофикация и загрязнение, для Северо-восточного – снижение прозрачности воды, эвтрофикация, загрязнение и изменение характера грунта, для Прикерченского района, собственно Азовского моря и лиманно-плавневой зоны – загрязнение и эвтрофикация.
Выводы
1. Видовое разнообразие морских фитоценозов экосистем Азово-Черноморского шельфа представлено 312 видами водных макрофитов, относящихся к 8 отделам: Cyanophyta (27 видов), Chlorophyta (65 видов), Charophyta (13 видов), Xanthophyta (1 вид), Rhodophyta (100 видов), Phaeophyta (47), Polypodiophyta (2 вида) и Magnoliophyta (57 видов). В Черном море макрофиты представлены 250 видами и 257 внутривидовыми таксонами как низших, так и высших растений, в Азовском море (включая лиманно-плавневые экосистемы) – 170 видами и 174 внутривидовыми таксонами. По мере продвижения от Черного моря к мелководным, опресненным лиманам с неустойчивым гидрологическим режимом снижается роль многолетних видов, при усилении роли эфемерных.
2. По сравнению с 60-ми годами ХХ века, в экосистемах российского шельфа Черного моря 66% всех видов водорослей-макрофитов не изменили своего статуса, 6% видов изменили статус с редкого и сопутствующего на сопутствующий и ведущий (это в основном зеленые водоросли), 23% видов изменили статус с сопутствующего и ведущего на редкий и сопутствующий (в основном красные водоросли) и 5% видов исчезли (в основном бурые водоросли).
3. Фитоценотическое разнообразие экосистем Азово-Черноморского шельфа представлено 54 ассоциациями, объединенными в 30 формаций. Доминирующей в шельфовых экосистемах Черного моря является формация Cystoseireta, Азовского моря, лиманов и плавней – Phragmiteta. Основная ассоциация Cystoseiretum dilophoso-cladostephosum имеет 3-4-х ярусное строение, простирается до глубины 10-15 м. Глубже (до 25-30 м) доминируют моно - и олигодоминантные 1-2-х ярусные ассоциации Codium vermilara и Phyllophora crispa. В антропогенно нарушенных акваториях первая ассоциация встречаются фрагментарно до глубины 10 м, а на глубинах 12-15 м растительность исчезает. Снижение прозрачности воды, ставшее причиной сокращения прибрежных зарослей, привело к полному исчезновению самых глубоководных ассоциаций (Chondrietum laurenciosum, Gracilarietum neriosum, Polysiphonietum subpurum, Antithamnietum subpurum) и сокращению ассоциаций средних этажей сублиторали (Cystoseiretum dilophoso-cladostephosum, Cystoseiretum phyllophorosum, Phyllophoretum purum).
4. Наиболее продуктивными в прибрежных экосистемах являются многолетние ассоциации бурых водорослей: самую высокую среднюю годовую биомассу и годовую продукцию имеют олигосапробные и α-мезосапробные ассоциации формации Cystoseireta: Cystoseiretum ulvosum, Cystoseiretum dilophosum и Cystoseiretum dilophoso-cladostephosum.
5. Наиболее уязвимыми в экосистемах российского шельфа Черного моря являются глубоководные фитоценозы и фитоценозы аккумулятивных ландшафтов, занятых сообществами морских трав, харовых водорослей, плохо переносящих эвтрофикацию. Сообщества твердых грунтов первых двух этажей сублиторали, более разнообразные по структуре и занимающие разные экологические ниши, более устойчивы к антропогенному воздействию. Наиболее широкой экологической валентностью обладают ассоциации формации Cystoseireta, имеющие достаточно мощный адаптационный потенциал, позволяющий им занимать акватории на глубинах от 0,1 до 10-15 м, различающихся степенью прибойности и сапробности.
6. Общая биомасса водных растений в юго-восточной части Азовского моря, включая лиманные и плавневые экосистемы превышает 2 млн. т. сырой массы, причем основную долю составляют пресноводные травы. Общая биомасса водорослей в Северо-восточном гидроботаническом районе Черного моря составляет около 185 тыс. т. сырой массы, из них 130 тыс. т. приходится на бурые водоросли рода цистозира. Юго-восточный и Анапско-Прикерченский гидроботанические районы Черного моря менее продуктивны – общая биомасса макрофитов составляет лишь около 8 и 5,5 тыс. т сырой массы соответственно.
7. За прошедшие 50 лет в экосистемах российского шельфа Черного моря произошли существенные изменения общей биомассы макрофитобентоса, обусловленные снижением прозрачности воды, эвтрофикацией и загрязнением шельфовой зоны. Полностью исчезли ассоциации зостеры, общие запасы цистозиры снизились более чем в 6 раз, запасы филлофоры – в 2 раза, а ульвы – несколько увеличились. Нижняя граница фитобентоса за последние полвека сместилась до глубины 32 м, что в 2-3 раза меньше, чем было ранее. Ущерб водным биоресурсам при нефтяном загрязнении акватории моря в районе морского терминала КТК, в 2003 г. составил 223 тонны сырой массы обоих видов цистозиры.
8. При усилении антропогенной нагрузки аборигенные фитоценозы сменяются ассоциациями эфемерных малопродуктивных видов, что сопровождается упрощением пространственной и иерархической структуры сообщества. При усилении загрязнения прибрежных экосистем нефтепродуктами происходят серьезные изменения в структуре морских фитоценозов. При достижении концентрации нефтепродуктов в донных отложениях 0,5 г/кг и сохранении ее в течение месяца (хроническое загрязнение) из экосистем шельфовой зоны исчезают популяции Сodium vermilara, образующие формацию Codieta на глубинах свыше 7-10 м Черного моря и популяции Zostera marina, занимающие мелководные акватории заливов Таманского полуострова и Керченского пролива. При концентрации нефтепродуктов выше 0,5 г/кг в течение указанного времени элиминируют эдификатор донных фитоценозов шельфа Черного моря Сystoseira crinita, а также Enteromorpha clathrata и Cladostephus spongiosus; при остром загрязнении (до 1 г/кг) элиминируют Сystoseira barbata, Phyllophora crispa, Gelidium spinosum и виды рода Laurencia. Устойчивые виды Cladophora albida, Ulva rigida, Callithamnion corrymbosum, Сeramium rubrum исчезают лишь в условиях аварийного загрязнения экосистемы при концентрации нефтепродуктов до 5 г/кг донных отложений.
9. У биоиндикаторного вида Cystoseira crinita произрастающей в эвтрофированной среде, наблюдается изменение отношения масс отделов ветвей в сторону развития адвентивного комплекса, снижение удельной поверхности слоевища. При увеличении комплексной загрязненности среды у цистозиры наблюдается снижение концентрации пигментов, активация ферментов антиоксидантной защиты. Виды, имеющие более высокое отношение каротиноидов к хлорофиллам и обладающие большей активностью антиоксидантных ферментов в норме, более устойчивы к загрязнению и эвтрофикации водной среды.
10. Результатом адаптации популяции цистозиры к эвтрофикации является омоложение, интенсификация метаболизма, уменьшение количества особей на единицу поверхности субстрата, что ведет к освобождению жизненного пространства для видов с большей функциональной активностью.
11. Для рационального использования запасов цистозиры без ущерба для популяций рекомендуется изъятие из прибрежных экосистем не более 5% биомассы в период с начала июня до начала ноября, с чередованием добычи по участкам, что позволит ежегодно изымать до 7 тыс. тонн сырой массы ценного природного сырья. Возможно восстановление популяций основных (в экологическом и хозяйственном отношении) видов - цистозиры, филлофоры, кодиума и других - с помощью гидробиотехнических конструкций, так как перечисленные виды входят во флору техногенных экотопов и могут формировать зрелые популяции на искусственных субстратах.
Список основных публикаций по теме диссертации
Статьи в журналах, рекомендованных ВАК
1. Макрофитобентос прибрежной акватории юго-восточной части Азовского моря // Защита окружающей среды в нефтегазовом комплексе. 2006. № 9. С. 116–119.
2. , , , Чередников токсичности среды обитания гидробионтов в районе лицензионного участка «Приазовнефть» методами биологического тестирования // Защита окружающей среды в нефтегазовом комплексе. 2006. № 9. С. 98–106.
3. Адаптационная пластичность морфологических и физиолого-биохимических параметров Cystoseira crinita // Известия ТИНРО. 2008. Том 152. С. 64–71.
4. Влияние нефтяного загрязнения на биохимические показатели Cystoseira crinita Bory // Труды Кубанского государственного аграрного университета. 2008. № 1 (10). С. 142–146.
5. Влияние нефтяного загрязнения донных отложений Азово-Черноморского бассейна на развитие Zostera marina L. // Труды Кубанского государственного аграрного университета. 2008. № 3 (12). С. 120–124.
6. Содержание некоторых фотосинтетических пигментов бурой водоросли Cystoseira barbata (Good. et Wood.) C. Ag. в условиях загрязнения донных отложений углеводородами нефти // Известия высших учебных заведений. Северо-Кавказский регион. Естественные науки. 2008. № 6. С. 60–63.
7. Запасы некоторых видов макрофитов на российском шельфе Черного моря: современное состояние, многолетняя динамика и анализ причин изменения // Известия ТИНРО. 2008. Том 155. С. 161–168.
8. , , , О проблеме оценки токсичности компонентов водной среды методами биологического тестирования // Вопросы рыболовства. 2008. Том 9. № 4 (36). С. 839–846.
9. , , Корпакова -функциональные особенности сообществ перифитона Темрюкского залива Азовского моря // Проблемы региональной экологии. 2009. № 4. С. 47–51.
10. Состав, структура и экологические особенности макрофитобентоса российской части Юго-восточного гидроботанического района Черного моря // Известия высших учебных заведений. Северо-Кавказский регион. Естественные науки. 2009. № 4. С. 64–67.
11. Оценка запасов и распределения некоторых видов макрофитов на российском шельфе Черного моря // Растительные ресурсы. 2009. Том 45. Вып. 3. С. 51–59.
Монографии и главы в коллективных монографиях
12. , Корпакова российского Азово-Черноморья. Ростов-на-Дону:
13. , , , , Громов водных и прибрежно-водных экосистем Азово-Черноморского бассейна. Краснодар: Просвещение-Юг. 20с.
14. , , Чередников Азовского моря: перифитон искусственных субстратов. Ростов-на-Дону:
15. , Степаньян -возрастная структура популяции цистозиры Северо-Кавказского побережья Черного и Азовского морей // Среда, биота и моделирование экологических процессов в Азовском море. Апатиты: изд. КНЦ РАН. 2001. С. 125–135.
16. , , Влияние различных видов загрязнения на морфобиохимические параметры макрофитобентоса // Среда, биота и моделирование экологических процессов в Азовском море. Апатиты: изд. КНЦ РАН. 2001. С. 195–218.
17. , Адаптивные возможности некоторых водорослей и морских трав к стрессовым условиям среды // Экосистемные исследования Азовского моря и побережья. Апатиты: изд. КНЦ РАН. 2002. С. 278–293.
18. , , Н, , Методы сбора и обработки гидробиологических проб // Методы рыбохозяйственных и природоохранных исследований в Азово-Черноморском бассейне. Краснодар: Просвещение-Юг. 2005. С. 50–79.
19. , , , , Цыбульская токсичности воды и донных отложений Керченского пролива и прилегающих акваторий Черного и Азовского морей по данным биотестирования // Керченская авария: последствия для водных экосистем. Ростов-на-Дону: –124.
Статьи в журналах и аналитических сборниках
20. , Экологические особенности бентосной растительности побережий Абрауского полуострова // Биоразнообразие полуострова Абрау: Сб. науч. тр. М.: изд-во МГУ. 2002. С. 30–37.
21. , , Фитобентос Таманского залива и Керченского пролива // Основные проблемы рыбного хозяйства и охрана рыбохозяйственных водоемов Азово-Черноморского бассейна: Сб. науч. тр. М.: «Национальные рыбные ресурсы». 2002. С. 170–176.
22. , , , Сушкова донных биоценозов в районе пос. Ю. Озерейка – Дюрсо // Основные проблемы рыбного хозяйства и охрана рыбохозяйственных водоемов Азово-Черноморского бассейна: Сб. науч. тр. Ростов н/Д: Эверест. 2004. С. 62–73.
23. Макрофитобентос российского сектора Черного моря // Экология моря. 2005. № 68. С. 19–25.
24. Функциональное состояние макрофитов в условиях экспериментального загрязнения донных отложений углеводородами нефти // Основные проблемы рыбного хозяйства и охраны рыбохозяйственных водоемов Азово-Черноморского бассейна: Сб. науч. тр. Ростов н/Д: Медиа-Пресс. 2006. С. 420–428.
25. , , , Барабашин проблемы оценки токсичности компонентов среды методами биотестирования // Защита окружающей среды в нефтегазовом комплексе. 2007. № 9. С. 23–29.
26. , , , Бычкова особенности лиманно-плавневой зоны Темрюкского залива Азовского моря // Защита окружающей среды в нефтегазовом комплексе. 2007. № 9. С. 69–75.
27. , , , , Барабашин токсичности среды обитания в юго-восточной части Азовского моря методами биологического тестирования // Защита окружающей среды в нефтегазовом комплексе. 2008. № 11. С. 62–69.
28. , , , , Бычкова характеристика юго-восточной части Азовского моря // Защита окружающей среды в нефтегазовом комплексе. 2008. № 11. С. 70–80.
29. , , , , Акинин рифовых систем для доочистки морской среды в Темрюкском заливе Азовского моря // Защита окружающей среды в нефтегазовом комплексе. 2009. № 11. С. 5–17.
30. , , Елецкий перифитона искусственных рифовых систем Темрюкского залива Азовского моря // Защита окружающей среды в нефтегазовом комплексе. 2009. № 11. С. 18–31.
Патенты
31. , , , Елецкий риф. Патент на изобретение № 000. Бюл. № 20 от 01.01.2001.
32. , , , Елецкий биотоп. Патент на изобретение № 000. Бюл. № 2 от 01.01.2001.
33. , , Аксенова определения токсичности контактной зоны «грунт – вода». Патент на изобретение № 000. Бюл. № 16 от 01.01.2001.
34. , , Чередников биотоп. Патент на изобретение № 000. Бюл. № 33 от 01.01.2001.
35. , , , Чередников биотоп. Патент на полезную модель № 000. Бюл. № 34 от 01.01.2001.
36. , , Корпакова для аэрации придонных слоев воды. Патент на полезную модель № 000. Бюл. № 26 от 01.01.2001.
37. , , , Чередников риф. Патент на полезную модель № 000. Бюл. № 28 от 01.01.2001.
38. , , , Инютина биотоп. Патент на полезную модель № 000. Бюл. № 26 от 01.01.2001.
Материалы докладов всероссийских и международных
конференций и съездов
39. , , Шевченко растительных сообществ Черного и Азовского море: оценка и перспективы использования // Проблемы сохранения экосистем и рационального использования биоресурсов Азово-Черноморского бассейна. Материалы международной научной конференции (8-12 октября 2001 г. Ростов-на-Дону). Ростов-на-Дону. 2001. С. 58–59.
40. , Степаньян возможности фитоценоза Cystoseira sp. к загрязнению и эвтрофии прибрежной зоны Черного моря // VIII съезд гидробиологического общества РАН (16-23 сентября 2001 г., Калининград). Калининград, 2001. Т. 2. С.102–103.
41. , Некоторые современные тенденции изменения макрофитобентоса Азово-Черноморского побережья // Сохранение и воспроизводство растительной компоненты биоразнообразия. Материалы международной конференции (28-31 мая 2002 г. Ростов-на-Дону). Ростов-на-Дону: РГУ. 2002. С. 81–84.
42. , , Прибрежные морские консорции и бентические пустыни // Эволюция морских экосистем под влиянием вселенцев и искусственной смертности фауны. Материалы международной научной конференции (12-15 июня 2003 г., Азов). Мурманск: ММБИ КНЦ РАН. 2003. С. 43–44.
43. , , Громов состояние водной растительности шельфа северокавказского побережья Черного моря // Ботанические исследования в азиатской России. Материалы XI съезда Русского ботанического общества (21-25 августа 2003г., Новосибирск-Барнаул). Т.1. Барнаул 2003. С. 71–72.
44. , Изменение морфо-функциональной и пространственной структуры донной растительности у кавказского побережья Черного моря // Биотехнология – охране окружающей среды. Научные труды международного биотехнологического центра МГУ. II международная научная конференция (25-27 мая 2004 г.). М.: Спорт и культура. 2004. С. 24.
45. Водоросли-макрофиты, как индикаторы загрязненности морских вод нефтепродуктами // Биотехнология – охране окружающей среды. Научные труды международного биотехнологического центра МГУ. II международная научная конференция (25-27 мая 2004 г.). М.: Спорт и культура. 2004. С. 95.
46. Afanasyev D. F. Present state of benthic macrophyte communities of North Caucasus coast of the Black sea // International workshop on the Black sea benthos. Istanbul. Published by Turkish Marine Research Foundation, Turkey. 2004. P. 153–157.
47. Донные фитоценозы северо-кавказского шельфа Черного моря и их продуктивность // Гидроботаника 2005. Материалы VI всероссийской школы-конференции по водным макрофитам (11-16 октября 2005 г., Борок). Рыбинск: Дом печати». 2006.-С. 204–207.
48. Продуктивность и запасы макрофитобентоса северо-кавказского побережья Черного моря // IX съезд гидробиологического общества РАН (18-22 сентября 2006 г., Тольятти). Тольятти. 2006. С. 23–24.
49. Влияние нефтепродуктов на некоторые биохимические показатели Cystoseira crinita Bory // Проблемы устойчивого функционирования водных и наземных экосистем. Материалы международной научной конференции (9-12 октября 2006 г., Ростов-на-Дону). Ростов-на-Дону. 2006. С. 20–21.
50. Afanasyev D. F. Macrophytobenthos productivity and its resources in the north-caucasian coastal area of the Black sea // Black sea ecosystem 2005 and beyond. 1st Biannual Scientific conference. Turkey, Istanbul Published by Commission on the protection of the Black sea against pollution, Turkey, 2006. – P. 106–109.
51. К анализу флоры водорослей-макрофитов Азово-Черноморского шельфа России // Фундаментальные и прикладные проблемы ботаники в начале XXI века. Материалы всероссийского съезда Русского ботанического общества (22-27 сентября 2008 г., Петрозаводск). Ч.2. Петрозаводск. 2008. С. 12–13.
52. , Громов водорослей-макрофитов Азовского моря и российского шельфа Черного моря // Современные проблемы альгологии. Материалы международной научной конференции и VII Школы по морской биологии (9-13 июня 2008 г. Ростов-на-Дону). Ростов-на-Дону: ЮНЦ РАН. 2008. С. 26–27.
53. Afanasyev D. F. Phytogeographical analysis of algal macrophytes of the Black sea Russian shelf // Climate change in the Black sea – hypothesis, observations, trends. Scenarios and mitigation strategy for the ecosystem. 2nd Biannual and Black sea scene EC project joint conference. Published by Commission on the protection of the Black sea against pollution, Bulgaria. 2008. P. 37–38.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
