


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Встречаемость генов inlF, inlG и inlH существенно отличалась от вышеописанных генов (табл. 5). Полимеразная цепная реакция давала положительный ответ только у части культур, что говорит о том, что у остальных культур L.monocytogenes данные гены отсутствуют, либо область связывания праймеров вариабельна.
Кроме того, проведенный скрининг показал вариабельность в размере гена inlF (табл.6). Аллель гена inlF, дающий укороченный продукт в ПЦР, преобладал среди культур, выделенных из морских гидробионтов. Данный аллель гена inlF не встречался среди культур L.monocytogenes из других источников.
Таблица 6
Вариабельность гена inlF у культур L. monocytogenes,
выделенных из различных источников
Продукт ПЦР с праймерами inlF1-inlF2 | Мертворожденные плоды (n=36), (% культур) | Животные (n=43), (% культур) | Продукты питания (n=27), (% культур) | |
Грызуны (n=19) | Морские гидробионты (n=24) | |||
нормальный | 100,0 | 84,2 | 4,2 | 51,8 |
укороченный | 0 | 0 | 70,8 | 0 |
удлиненный | 0 | 0 | 4,2 | 3,7 |
отсутствует | 0 | 15,8 | 20,8 | 44,4 |
Экспериментальное изучение рядом авторов роли продуктов генов inlG и inlH показало, что эти белки не только не являются необходимыми в процессе внутриклеточной инфекции, но, напротив, частично подавляют адгезию и инвазию L. monocytogenes in vitro (Bergmann B. et al., 2002). В наших исследованиях эти гены были выявлены у двух из трех культур L.monocytogenes, относящихся ко второй филогенетической линии и изолированных у здоровых носителей, в то время как среди культур, выделенных от больных людей с клиническими проявлениями листериоза, ген inlG был выявлен у двух, а inlH у одного из пяти изолятов. Важно также отметить, что гены inlG и inlH не выявлялись у подавляющего большинства штаммов L. monocytogenes, изолированных из животных. Полученные нами данные подтверждают предположение о том, что белки InlG и InlH не являются необходимыми для развития листериозной инфекции.
Таким образом, установлено, что штаммоспецифический полиморфизм спектра интерналинов у L.monocytogenes коррелирует с источником выделения штамма и не зависит от региона изоляции.
Результаты, полученные при выявлении генов, кодирующих белки ActA и Auto, показали наличие гена, кодирующего белок ActА, у всех без исключения штаммов L.monocytogenes. Ген аut, кодирующий белок Auto, методом ПЦР обнаруживали менее чем у половины штаммов L.monocytogenes, изолированных как на Дальнем Востоке, так и в Европейской части России (32,9 и 25% соответственно). Ген аut, чаще определялся среди L.monocytogenes, изолированных из пищевых продуктов (51,6 %), и реже - среди штаммов листерий, изолированных из клинического материала (34,5 %) и из органов грызунов (18,7 %).
Таким образом, нами были выявлены гены факторов инвазии, которые встречаются у подавляющего большинства изолятов L.monocytoge-nes,выделенных из эукариотических организмов, включая человека, - inlA, inlB, inlC, inlE. Эти гены были использованы нами в качестве мишеней для разработки системы генотипирования.
Сравнительный анализ полиморфизма генов, кодирующих белки-интерналины (inlA, inlB, inlC, inlE), и гена prs у штаммов L.monocytogenes, изолированных в Дальневосточном регионе и Европейской части России
Анализ полиморфизма был проведен для генов факторов инвазии, которые, как описано выше, присутствовали у всех культур L. monocytogenes. Такими генами являются гены интерналинов inlA, inlB, inlC, inlE. Была изучена вариабельность внутренних фрагментов этих генов, а именно фрагментов, кодирующих LRR-домены, непосредственно вовлеченные в процесс взаимодействия интерналинов со специфическими рецепторами эукариот. Кроме генов, кодирующих интерналины, был секвенирован внутренний фрагмент гена prs, кодирующего фосфорибозилсинтазу - белок общего метаболизма, выявленный у всех представителей рода Listeria (табл.7).
Таблица 7
Сравнительный анализ секвенированных фрагментов генов у культур L.monocytogenes, изолированных в Дальневосточном регионе и Европейской части Российской Федерации
Ген | Длина фрагмента, пн (% общей ОРФ) | Амино-кислоты | Число замен | Число аллелей | ||||
синони-мичных | несинонимичных | линия специфичных несинони-мичных | общее | специ-фичных для ЕЧ | специ-фич-ных для ДВ | |||
inlA | 54-269 | 14 | 6 | 0 | 11 | 5 | 3 | |
inlB | 64-269 | 37 | 25 | 7 | 18 | 5 | 12 | |
inlC | 70-264 | 17 | 8 | 4 | 12 | 3 | 7 | |
inlE | 82-267 | 45 | 46 | 29 | 11 | 4 | 4 | |
prs | 39-244 | 24 | 1 | 0 | 6 | 3 | 1 |
Примечание: ОРФ - открытая рамка считывания, ЕЧ – Европейская часть России, ДВ – Дальний Восток.
У гена prs все замены (24), кроме одной, были синонимичны. Отсутствие несинонимичных замен в последовательности гена prs подтверждает представление об отсутствии адаптивной вариабельности среди белков общего метаболизма и отражает аминокислотную стабильность этого белка для изолятов L.monocytogenes, эволюционирующих в разных условиях.
Напротив, интерналины характеризовались значительной изменчивостью, в том числе на аминокислотном уровне. Наибольшую вариабельность продемонстрировали гены inlE и inlB, у которых было выявлено 45 и 37 синонимичных, 46 и 25 несинонимичных замен, соответственно. Фрагменты генов inlA и inlC были более консервативны (14 и 17 – синонимичных, 6 и 8 – несинонимичных замен соответственно). Установлено наличие специфических аллелей генов inlA, inlB, inlC, inlE, prs, присущих определенному географическому региону (табл.7).
Для всех пяти маркерных фрагментов в сумме было выявлено 223 единичных нуклеотидных замен. Высокая вариабельность изученных генов позволила нам разработать систему типирования, описанную ниже.
Филогенетическая характеристика популяции L.monocytogenes, распространенной на Дальнем Востоке Российской Федерации
Прежде чем приступать к разработке независимой системы типирования, мы охарактеризовали филогенетическую структуру популяции L.monocytogenes, распространенной на Дальнем Востоке, уже существующими методами.
Известно, что бактерии вида L.monocytogenes имеют сложную антигенную структуру и включают 13 серологических вариантов. Большая часть случаев заболеваний листериозом связана с серовариантами 4b, 1/2a, 1/2b, причем эпидемические вспышки заболевания чаще всего связаны с серовариантом 4b. За последние годы установлено, что внутри вида L.monocytogenes выделяют три филогенетические линии, отличающиеся по своему эпидемиологическому потенциалу, каждая из которых объединяет штаммы, принадлежащие к определенным серовариантам (Piffaretti J. C. et al., 1989, Wiedman M. et al., 1997, Doumith M. et al., 2004).
С целью оценки вариабельности и эпидемиологической значимости изолятов L.monocytogenes, выделенных из разнообразных источников, первоначально нами были использованы методы серо - и фаготипирования.
В нашей стране для серотипирования используются сыворотки двух вариантов. Отмечено, что большинство дальневосточных культур L.monocytogenes, принадлежали ко второй серогруппе согласно отечественной классификации. Культуры L.monocytogenes, изолированные из пищевых продуктов, в большинстве (72,2±7,5%) относились к первой серогруппе (рис.1).

Рис. 1. Частота встречаемости (%) L.monocytogenes различных серогрупп на территории РФ: ДВ – Дальний Восток, ЕЧ – Европейская часть РФ.
Результаты, полученные при изучении коллекции L.monocytogenes, показали, что среди дальневосточных L. monocytogenes 34,1±7,1% культур типировались бактериофагом L-4A, 15,9±5,5 % культур – бактериофагом L-2А (рис.2).
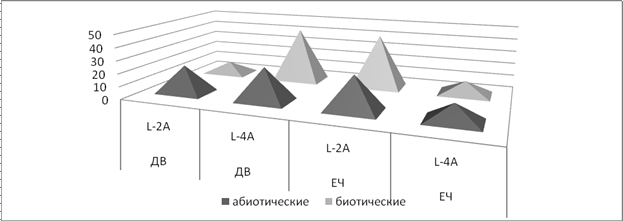
Рис. 2. Фаговарианты (%) культур L.monocytogenes, изолированных из различных источников в Дальневосточном регионе (ДВ) и Европейской части (ЕЧ) России.
В то же время анализ антигенной структуры штаммов L.monocytogenes с использованием отечественных сывороток и бактериофагов двух типов показал относительно низкую специфичность данной системы дифференциации. Кроме того, ее низкая разрешающая способность не позволяет проводить анализ эпидемически значимых штаммов листерий.
Исследования, выполненные с помощью метода мультиплексной ПЦР, предложенной (Doumith М. al., 2004), позволили более детально охарактеризовать структуру популяции L.monocytogenes, распространенной на разных территориях России, и прежде всего, установить преобладание эпидемиологически значимого сероварианта 4b среди L. monocytogenes, выделенных на Дальнем Востоке (табл. 8).
Таблица 8
Типирование культур L.monocytogenes,изолированных в
Дальневосточном регионе и Европейской части РФ, на основе выявления генетических маркеров, специфичных для определенных серогрупп
Серогруппа | Частота выделения культур в зависимости от территории изоляции L.monocytogenes (%) | p | |
Дальний Восток (n=52) | Европейская часть (n=17) | ||
1/2a | 19,2±5,5 | 41,17±12,3 | < 0,05 |
1/2b | 3,8±2,6 | 11,8±8,1 | > 0,05 |
1/2c | 1,92±1,9 | 17,64±9,5 | < 0,05 |
4b | 75,4±6,0 | 29,4±11,4 | < 0,01 |
Таким образом, полученные результаты показали гетерогенность штаммов L.monocytogenes, изолированных из различных объектов на территории России. Однако использованные методы характеризовались низкой дискриминационной способностью, не отражали полной картины вариабельности популяции и не выявляли степени генетического родства между эпидемическими и неэпидемическими штаммами L.monocytogenes.
В последние годы для типирования наиболее значимых в эпидемиологическом отношении штаммов L.monocytogenes разрабатываются молекулярно-генетические методы, среди которых мультилокусное секвенирование превосходит все остальные по воспроизводимости и, во многих случаях, по разрешающей способности. Для разработки системы типирования нами было выбрано 40 независимых культур L. monocytogenes, изолированных в разных регионах России (Дальний Восток и Европейская часть России), у которых было осуществлено секвенирование внутренних областей генов inlA, inlB, inlC, inlE и prs, по схеме, описанной выше (табл. 7).
Суммарная длина конкатемеров, включающих фрагменты 5 маркерных генов, равнялась 3029 н. п., что составляет около 0,1% хромосомной ДНК L.monocytogenes. Изучаемые последовательности относились к 25 аллелям. Только один изолят из 40 имел продолжительную делецию – изолят VIMHA009 имел делецию 69 п. н. в гене inlE. Остальные различия заключались в единичных нуклеотидных заменах. Вычисление индекса дискриминации (D. I.) для предлагаемого метода типирования дало величину 0,982. Это означает, что если два эпидемически не связанных изолята, принадлежащих к разным штаммам, будут произвольно выбраны из популяции, то в предлагаемом методе типирования с вероятностью 98 % они будут относиться к разным типам. Полученная величина, хотя и не является максимально возможной (теоретически максимально возможная величина для индекса дискриминации равна 1), обеспечивает приемлемый уровень дифференциации эпидемически не связанных изолятов.
Для установления филогенетических связей между изолятами L.monocytogenes была построена дендрограмма, с использованием метода минимальной эволюции (рис.3), которая позволила разделить L.monocytogenes на две группы, соответствующие описанным филогенетическим линиям и коррелирующие с принадлежностью к определенному сероварианту ( и соавт., 2003, Piffaretti J. C. et al., 1989, Wiedman M. et al., 1997).
Большинство культур L.monocytogenes, изолированных в Дальневосточном регионе, принадлежали к первой филогенетической линии, наиболее опасной в эпидемическом плане (по Wiedman M. et al., 1997). Было отмечено, что среди изолятов L. monocytogenes, полученных из клинического материала (от мертворожденных плодов), от грызунов и морских гидробионтов преобладают штаммы листерий, относящиеся к первой филогенетической линии; среди изолятов L. monocytogenes из продуктов питания доминируют штаммы, относящиеся ко второй филогенетической линии.

Рис. 3. Дендрограмма, устанавливающая степень схожести последовательностей ДНК среди изолятов L.monocytogenes.
Овальные скобки – два основных кластера, соответствующих I (нижняя) и II (верхняя) клональным линиям. Окружности – группы изолятов, имеющих идентичные последовательности изученных фрагментов; * – изоляты из разных регионов, образующие совместные кластеры.
Таким образом, в результате проведенных исследований нами впервые установлена филогенетическая структура L. monocytogenes, распространенных на Дальнем Востоке, а также подтверждено существование двух филогенетических линий внутри вида L.monocytogenes.
Анализ филогенетической принадлежности изолятов L.monocytogenes в зависимости от региона и источника выделения
Разработанная система была применена для анализа распределения штаммов L. monocytogenes в зависимости от региона и источника выделения. Используя данную систему, была осуществлена дифференциация всех имеющихся изолятов L.monocytogenes, выделенных в Дальневосточном и Европейском регионах РФ, которая выявила 32 сиквенс-типа (СТ) (табл.9).
Отмечено, что все сиквенс-типы имели региональную приуроченность. Среди изолятов листерий, выделенных на Дальнем Востоке, было найдено 19, а в Европейской части России – 13 сиквенс-типов. Не было найдено ни одного сиквенс-типа, который встречался бы в обоих регионах. Среди дальневосточных L.monocytogenes заметно преобладали СТ 1, СТ 9 и СТ 17. Не отмечено превалирования какого-либо сиквенс-типа среди L.monocytogenes, изолированных в Европейской части РФ.
Таким образом, доказано существование регион-специфического распределения штаммов L.monocytogenes на территории РФ. Кроме того, выявленное разнообразие сиквенс-типов предполагает поликлональное распределение вида L. monocytogenes в пределах географически отдаленных областей.
При анализе распределения сиквенс-типов в зависимости от источника выделения, было найдено, что некоторые сиквенс-типы обнаруживаются среди специфических хозяев с более высокой частотой, по сравнению с другими сиквенс-типами: СТ1, СТ5, СТ9 и СТ17 чаще встречались среди изолятов L.monocytogenes, полученных из мертворожденных плодов (группа «МП»), СТ1 – среди изолятов группы «Г» и СТ17 в группе «МГ» (табл. 9).
Таблица 9
Характеристика сиквенс-типов (СТ) среди изолятов L.monocytogenes,
полученных на Дальнем Востоке и в Европейской части РФ
СТ | Регион | Аллель | Число изолятов | |||||||||
Общее | В группе специфичного хозяина | |||||||||||
inlA | inlB | inlC | inlE | prs | «МП» | «Н» | «Г» | «МГ» | «П» | |||
1 | ДВ | 1 | 1 | 1 | 1 | 1 | 12 | 4 | 0 | 7 | 0 | 1 |
2 | ДВ | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
3 | ДВ | 1 | 9 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
4 | ДВ | 1 | 9 | 1 | 4 | 2 | 2 | 1 | 0 | 0 | 0 | 1 |
5 | ДВ | 1 | 9 | 1 | 3 | 2 | 4 | 4 | 0 | 0 | 0 | 0 |
6 | ДВ | 1 | 10 | 1 | 3 | 2 | 1 | 1 | 0 | 0 | 0 | 0 |
7 | ЕЧ | 5 | 14 | 6 | 8 | 4 | 1 | 1 | 0 | 0 | 0 | 0 |
8 | ЕЧ | 5 | 14 | 6 | 10 | 5 | 1 | 1 | 0 | 0 | 0 | 0 |
9 | ДВ | 9 | 14 | 9 | 6 | 5 | 7 | 3 | 2 | 0 | 1 | 1 |
10 | ЕЧ | 8 | 14 | 6 | 10 | 5 | 2 | 0 | 1 | 0 | 0 | 1 |
11 | ДВ | 2 | 1 | 5 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 |
12 | ДВ | 10 | 14 | 8 | 7 | 6 | 2 | 0 | 0 | 2 | 0 | 0 |
13 | ЕЧ | 9 | 14 | 6 | 6 | 5 | 1 | 0 | 0 | 1 | 0 | 0 |
14 | ЕЧ | 2 | 7 | 4 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
15 | ЕЧ | 4 | 14 | 6 | 8 | 4 | 4 | 0 | 0 | 1 | 0 | 3 |
16 | ЕЧ | 6 | 12 | 6 | 6 | 5 | 1 | 0 | 0 | 0 | 0 | 1 |
17 | ДВ | 2 | 1 | 6 | 1 | 1 | 10 | 3 | 0 | 0 | 9 | 1 |
18 | ДВ | 2 | 5 | 4 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
19 | ДВ | 2 | 6 | 4 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
20 | ДВ | 2 | 2 | 4 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
21 | ДВ | 3 | 8 | 3 | 1 | 2 | 1 | 0 | 1 | 0 | 0 | 0 |
22 | ДВ | 2 | 1 | 4 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 |
23 | ЕЧ | 8 | 15 | 6 | 10 | 5 | 1 | 0 | 1 | 0 | 0 | 0 |
24 | ЕЧ | 4 | 15 | 6 | 8 | 4 | 1 | 0 | 1 | 0 | 0 | 0 |
25 | ЕЧ | 1 | 16 | 2 | 3 | 3 | 1 | 0 | 1 | 0 | 0 | 0 |
26 | ЕЧ | 4 | 14 | 6 | 8 | 4 | 1 | 0 | 1 | 0 | 0 | 0 |
27 | ЕЧ | 11 | 14 | 11 | 11 | 5 | 1 | 0 | 1 | 0 | 0 | 0 |
28 | ЕЧ | 9 | 11 | 7 | 8 | 6 | 1 | 0 | 0 | 0 | 0 | 1 |
29 | ДВ | 1 | 4 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
30 | ДВ | 7 | 18 | 9 | 6 | 5 | 1 | 0 | 0 | 0 | 0 | 1 |
31 | ДВ | 10 | 17 | 12 | 2 | 5 | 1 | 0 | 0 | 0 | 0 | 1 |
32 | ДВ | 7 | 13 | 10 | 9 | 5 | 1 | 0 | 0 | 0 | 0 | 1 |
Примечание: ДВ – Дальний Восток, ЕЧ – Европейская часть России, «МП» – изоляты L.monocytogenes, выделенные из мертворожденных плодов, «Н» – изоляты от носителей, «Г» - изоляты от грызунов, «МГ» – изоляты из морских гидробионтов, «П» – изоляты из продуктов, объектов окружающей среды и млекопитающих (кроме грызунов).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
