
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
аналогов Spm и Spd с ферментами метаболизма полиаминов и клетками [†]
Spm, Spd и их N1-ацетильные производные представляют собой ахиральные молекулы и до начала наших исследований в литературе отсутствовали какие-либо данные о стереоспе-цифичности ферментов их метаболизма. Вместе с тем, исходя из общих соображений, а так-же основываясь на различиях в активности хиральных аналогов дезоксиспергуалина (Lebre-ton L., et al. 1999. J.Med.Chem., 42, 4749–4763) и гидроксилированных производных диэтил-нор-спермина (Bergeron R. J., et al. 2001. J. Med. Chem., 44, ), можно было предполо-жить существование скрытой стереоспецифичности соответствующих ферментов. Если это предположение справедливо, то использование оптических изомеров a-метилированных ана-логов Spm и Spd открывает неизвестные ранее подходы к тонкому регулированию активнос-ти ферментов метаболизма полиаминов и, что не менее важно, новые возможности преци-зионного воздействия на гомеостаз полиаминов в клетке.
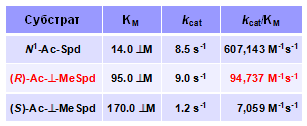 Уже первые эксперименты показали, что флавин-зависимая ацетилполиаминооксида-за (АРАО) предпочтительно окисляет (R)-изомер N-Ac-a-MeSpd (Табл. 1).Таким образом, впервые было показано, что АРАО, природными субстратами которой служат ахиральные N1-Ac-Spd и N1-Ac-Spm, обладает скрытой сте-реоспецифичностью.
Уже первые эксперименты показали, что флавин-зависимая ацетилполиаминооксида-за (АРАО) предпочтительно окисляет (R)-изомер N-Ac-a-MeSpd (Табл. 1).Таким образом, впервые было показано, что АРАО, природными субстратами которой служат ахиральные N1-Ac-Spd и N1-Ac-Spm, обладает скрытой сте-реоспецифичностью.
Табл. 1. Окисление расщепление N1-Ac-Spd и изоме-ров N-Ac-a-MeSpd под действием (AРАО).
Известно, что АРАО способна в присутствии ароматических альдегидов эффективно окис-лять и неацетилированный Spd (Holtta E. 1977. Biochemistry, 16, 91–100). Механизм этой реак-ции не исследован, а первоначальное предположение о промежуточном образовании в раство-ре оснований Шиффа (Holtta E. 1977. Biochemistry, 16, 91–100) представляется маловероятным при проведении реакции в 0.1 М глициновом буфере, а также, учитывая что взаимодействие ароматических альдегидов с 1,3-диаминопропанами приводит к достаточно сложной смеси продуктов, включая и циклические структуры (Metzler C. М., et al. 1980. J.Am.Chem.Soc., 102, ). Оказалось, что бензальдегид (ВА) катализирует расщепление преимущественно (R)-a-MeSpd, а в присутствии пиридоксаля (PL) субстратом оказывается уже (S)-изомер (Табл. 2). Напротив, 4-пиридинальдегид (4-Py-Ald) неспособен осуществлять стереоконтроль реак-ции и одинаково эффективен как в случае (R)-, так и (S)-изомеров a-MeSpd
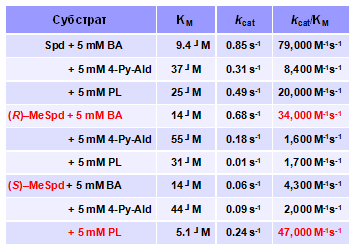 Табл. 2. Окислительное расщепление Spd и изомеров a-MeSpd AРАО. в присутствии бенз-альдегида (ВА), пиридоксаля (PL) и 4-пири-динальдегида (4-Py-Ald).
Табл. 2. Окислительное расщепление Spd и изомеров a-MeSpd AРАО. в присутствии бенз-альдегида (ВА), пиридоксаля (PL) и 4-пири-динальдегида (4-Py-Ald).
Таким образом, при помощи ахиральных ароматических альдегидов нам впервые удалось найти изящный способ регуля-ции стереоспецифичности АРАО, позво-ляющий избирательно окислять аналоги субстрата только (R)- или только (S)-конфигурации, что весьма необычно не только для флавин-зависимых оксидаз, но и для энзимологии в целом.
Недавно в животных клетках была обнаружена сперминоксидаза (SMO) – новый фла-вин-зависимый фермент, окисляющий Spm в Spd без предварительного N1-ацетилирования (Wang Y., et al. 2001. Cancer Res., 61, ). Так как родственная флавин-зависимая AРАО обладает скрытой стереоспецифичностью (см. выше), то мы исследовали взаимодейст-вие (R,R)- и (S,S)-диастереомеров a, a’-Me2Spm с рекомбинантной SMO. Оказалось, что суб-стратными свойствами обладал лишь (S,S)- a, a’-Me2Spm, а эффективность его расщепления была даже несколько выше, чем у Spm (Табл. 3). Таким образом, нам впервые удалось пока - казать, что SMO, как и АРАО, обладает 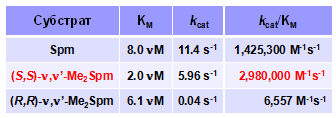 скрытой стереоспецифичностью. При этом, АРАО предпочтительно расщепляет (R)-N-Ac-a-MeSpd, а субстратом SMO, напротив, служит лишь (S,S)-a, a’-Me2Spm.
скрытой стереоспецифичностью. При этом, АРАО предпочтительно расщепляет (R)-N-Ac-a-MeSpd, а субстратом SMO, напротив, служит лишь (S,S)-a, a’-Me2Spm.
Табл. 3. Окислительное расщепление диастерео-меров a,a’-Me2Spm сперминоксидазой.
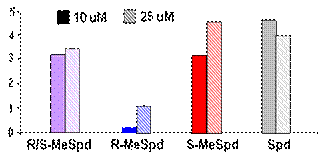
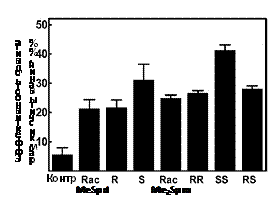
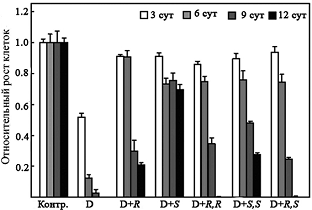 Аминокислота гипузин (Ne-(4-амино-2-оксибутил)-L-лизин), входящая в состав высо-коконсервативного участка фактора инициации трансляции eIF5A, присутствующего у всех эукариот, представляет собой одно из наиболее известных и метаболически-значимых кова-лентных производных Put (см. обзор: Park M. H. 2006. J.Biochem.(Tokyo), 139, 161–169). Так как немодифицированный eIF5A неактивен, то не всегда возможно однозначно определить, связано ли замедление роста клеток, вызванное снижением внутриклеточной концентрации Spd (единственный донор аминобутильного фрагмента), только с нарушением биосинтеза ги-пузина или имеет другие полиамин-зависимые причины. Располагая системой изомеров a-MeSpd и диастереомеров a, a’-Me2Spm, мы попытались дискриминировать гипузин-опосре-дованное ингибирование роста клеток и последствия, прямо не связанные с нарушением био-синтеза гипузина. Оказалось, что (R)- и (S)-изомеры a-MeSpd, а также и все три диастереоме-ра a, a’-Me2Spm, в равной степени способны поддерживать рост клеток DU145 в течение первых 6 сут инкубации с DFMO (Рис. 3). Однако при более продолжительном времени ин-гибирования ODC снижение внутриклеточной концентрации Spd становится критическим.
Аминокислота гипузин (Ne-(4-амино-2-оксибутил)-L-лизин), входящая в состав высо-коконсервативного участка фактора инициации трансляции eIF5A, присутствующего у всех эукариот, представляет собой одно из наиболее известных и метаболически-значимых кова-лентных производных Put (см. обзор: Park M. H. 2006. J.Biochem.(Tokyo), 139, 161–169). Так как немодифицированный eIF5A неактивен, то не всегда возможно однозначно определить, связано ли замедление роста клеток, вызванное снижением внутриклеточной концентрации Spd (единственный донор аминобутильного фрагмента), только с нарушением биосинтеза ги-пузина или имеет другие полиамин-зависимые причины. Располагая системой изомеров a-MeSpd и диастереомеров a, a’-Me2Spm, мы попытались дискриминировать гипузин-опосре-дованное ингибирование роста клеток и последствия, прямо не связанные с нарушением био-синтеза гипузина. Оказалось, что (R)- и (S)-изомеры a-MeSpd, а также и все три диастереоме-ра a, a’-Me2Spm, в равной степени способны поддерживать рост клеток DU145 в течение первых 6 сут инкубации с DFMO (Рис. 3). Однако при более продолжительном времени ин-гибирования ODC снижение внутриклеточной концентрации Spd становится критическим.
Рис. 3. Изомеры a-MeSpd и диастереомеры a,a’-Me2Spd по-разному поддерживают рост клеток DU145 с истощенным пулом Spd и Put. Клетки DU145 выращивали в присутствии 5 mM DFMO в течение 3, 6, 9, или 12 дней без добавления a-MeSpd/ a, a’-Me2Spm (контроль), или в присутствии аналогов (100 mM). D, DFMO; R, (R)-a-MeSpd; S, (S)-a-MeSpd; RR, (R,R)-a, a’-Me2Spm; SS, (S,S)-a, a’-Me2Spm; RS, (R,S)- a, a’-Me2Spm
Из исследованных аналогов лишь (S)-a-MeSpd и, в меньшей степени, (S,S)-a, a’-Me2Spm, который частично превращался в (S)-a-MeSpd, эффективно поддерживали рост клеток. Эти результаты прямо указывают на существование двух независимых причин замедления роста клеток, вызываемых истощением пула Spd.
Неспособность (R)-a-MeSpd поддерживать рост клеток DU145 при инкубации с DFMO более 6 сут привела к необходимости исследования субстратных свойств (R)- и (S)-изомеров a-MeSpd в дезоксигипузинсинтазной реакции, которая является скорость-опреде-ляющей стадией биосинтеза гипузинилированного eIF5A (Рис. 4). Субстратные свойства изо-
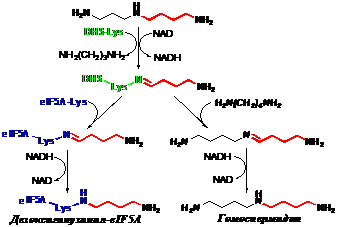 меров оценивали по образованию гомоспермидина, который в отсутствии eIF5A возникает при инкубации дезоксигипузинсинтазы (DHS) cо Spd или его аналогами в присутствии аль-тернативного акцептора аминобутильного фрагмента – [14С]-Put.
меров оценивали по образованию гомоспермидина, который в отсутствии eIF5A возникает при инкубации дезоксигипузинсинтазы (DHS) cо Spd или его аналогами в присутствии аль-тернативного акцептора аминобутильного фрагмента – [14С]-Put.
Рис. 4. Образования дезоксигипузинил-eIF5A и гомоспермидина в дезоксигипузинсинтазной реакции (по данным работы: Park J-H., et al. 2003. J.Biol.Chem., 278, )
Донором аминобутильного фрагмента в де-зоксигипузинсинтазной реакции оказался лишь (S)-a-MeSpd, т. е. впервые было обна-ружено, что дезоксигипузинсинтаза облада-ет скрытой стереоспецифичностью (Рис. 5). Это свойство фермента позволило при помощи
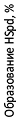
 (R)- и (S)-изомеров a-MeSpd показать, что сначала замедление роста клеток определяется де-фицитом собственно полиаминов, а гипузин-опосредованные эффекты начинают проявлять-ся существенно позднее.
(R)- и (S)-изомеров a-MeSpd показать, что сначала замедление роста клеток определяется де-фицитом собственно полиаминов, а гипузин-опосредованные эффекты начинают проявлять-ся существенно позднее.
Рис. 5. Субстратные свойства стереоизомров
a-MeSpd в дезоксигипузинсинтазной реакции. Реакционная смесь (50 ml) содержала 0.2 M Gly-NaOH буфер (pH, 9.5), 50 mg BSA, 1 mM DTT, 1 mCi [14C]-Put (100 mCi/mmol), 1 mM NAD, 10 mM и 25 mM Spd или 10 mM и 25 mM изомеров MeSpd и 1.0 mg рекомбинантной DHS человека.
Системе активного транспорта Spm и Spd отводится важное место в поддержании го-меостаза полиаминов. Поэтому были исследованы особенности транспорта хиральных аналогов Spm и Spd в клетки DU145. Оказалось, что со-держание изомеров a-MeSpd и диастереомеров a, a’-Me2Spm в клетках было сопоставимо с содержанием Spm и Spd, что указывало на активный транспорт аналогов (данные не приведе-ны). Однако изомеры a-MeSpd и диастереомеров a, a’-Me2Spm по-разному конкурировали за транспорт со Spd и Spm, а наименее эффективными оказались (S)-a-MeSpd и (S,S)-a, a’-Me2Spm (Рис. 6).
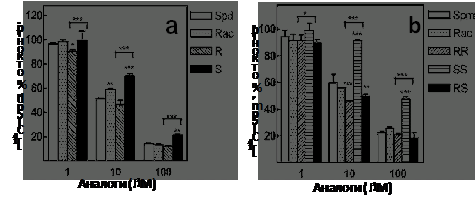 Рис. 6. Транспорт стререо-изомеров MeSpd (a) и Me2Spm (b) в клетки DU145. Клетки инкубиро-вали в течение 10 мин с 10 mM [14C]-Spd (a) или 10 mM [14C]-Spm (b) в присутствии 1, 10 или 100 mM исследуе-мого стереоизомера. R, (R)-a-MeSpd; S, (S)-a-MeSpd; RR, (R,R)-a, a’-Me2Spm; SS, (S,S)-a, a’-Me2Spm; RS, (R,S) -a, a’-Me2Spm.
Рис. 6. Транспорт стререо-изомеров MeSpd (a) и Me2Spm (b) в клетки DU145. Клетки инкубиро-вали в течение 10 мин с 10 mM [14C]-Spd (a) или 10 mM [14C]-Spm (b) в присутствии 1, 10 или 100 mM исследуе-мого стереоизомера. R, (R)-a-MeSpd; S, (S)-a-MeSpd; RR, (R,R)-a, a’-Me2Spm; SS, (S,S)-a, a’-Me2Spm; RS, (R,S) -a, a’-Me2Spm.
Таким образом, система транспорта полиаминов обладает стереоспецифичностью, что позво-
ляет регулировать доступность их аналогов, изменяя конфигурацию хирального центра.
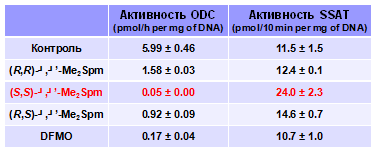 Одним из важнейших регуляторов внутриклеточного содержания полиаминов являет-ся небольшой белок антизим (AZ), биосинтез которого индуцируется в ответ на повышение концентрации полиаминов в клетке (Spm и Spd вызывают +1 сдвиг рамки считывания мРНК AZ, приводящий к образованию активного белка). AZ связывается с ODC, вызывая диссоциа-цию фермента на субъединицы, которые не обладают декарбоксилазной активностью, что приводит к снижению активности ODC в клетке. Однако эффекты диастереомеров a, a’-Me2Spm значительно отличались, а (S,S)-a, a’-Me2Spm по своей активности превосходил даже DFMO (Табл. 4).
Одним из важнейших регуляторов внутриклеточного содержания полиаминов являет-ся небольшой белок антизим (AZ), биосинтез которого индуцируется в ответ на повышение концентрации полиаминов в клетке (Spm и Spd вызывают +1 сдвиг рамки считывания мРНК AZ, приводящий к образованию активного белка). AZ связывается с ODC, вызывая диссоциа-цию фермента на субъединицы, которые не обладают декарбоксилазной активностью, что приводит к снижению активности ODC в клетке. Однако эффекты диастереомеров a, a’-Me2Spm значительно отличались, а (S,S)-a, a’-Me2Spm по своей активности превосходил даже DFMO (Табл. 4).
Табл. 4. Влияние диастереомеров a,a’-Me2Spm (100 mM) на активности ODC и SSAT через 72 ч инкубации клеток DU145 с с аналогами Spm.
Таким образом, изменяя конфигура-цию хирального центра a-MeSpd (дан-ные не приведены) и a, a’-Me2Spm, удается регулировать активность ODC в клетке, по-види-мому, через AZ-опосредованные механизмы.
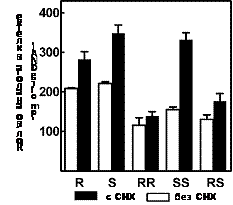 Известно, что помимо диссоциации ODC на субъединицы AZ ингибирует и транспорт полиаминов в клетки. Поэтому был исследован транспорт изомеров a-MeSpd и a, a’-Me2Spm в присутствии и отсутствии СНХ (ингибитор биосинтеза белка в клетке). Оказалось, что ин-гибирование биосинтеза AZ сильнее всего сказывается на накоплении (S,S)-a, a’-Me2Spm и (S)-a-MeSpd (Рис. 7), т. е. именно тех изомеров, которые наиболее эффективно сни-жают активность ODC в клетке (Табл. 4).
Известно, что помимо диссоциации ODC на субъединицы AZ ингибирует и транспорт полиаминов в клетки. Поэтому был исследован транспорт изомеров a-MeSpd и a, a’-Me2Spm в присутствии и отсутствии СНХ (ингибитор биосинтеза белка в клетке). Оказалось, что ин-гибирование биосинтеза AZ сильнее всего сказывается на накоплении (S,S)-a, a’-Me2Spm и (S)-a-MeSpd (Рис. 7), т. е. именно тех изомеров, которые наиболее эффективно сни-жают активность ODC в клетке (Табл. 4).
Рис. 7. Влияние биосинтеза AZ на транспорт стереоизомеров MeSpd и Me2Spm в клетки DU145. Клетки инкубировали с или без CHX (10 mg/ml) в течение 1 ч, а затем в присутствии аналогов (100 mM) еще в течение 2 ч. R, (R)-a-MeSpd; S, (S)-a-MeSpd; RR, (R,R)-a, a’-Me2Spm; SS, (S,S)-a, a’-Me2Spm; RS, (R,S)- a, a’-Me2Spm.
 Таким образом, гипотеза о возможности стереоконтроля +1 сдвига рамки считывания мРНК AZ получила дополнительные подтверждения. В заклю-чение было исследовано влияние стереоизомеров a-MeSpd и a, a’-Me2Spm на собственно +1 сдвиг рамки считывания мРНК (Рис. 8). Оказалось,
Таким образом, гипотеза о возможности стереоконтроля +1 сдвига рамки считывания мРНК AZ получила дополнительные подтверждения. В заклю-чение было исследовано влияние стереоизомеров a-MeSpd и a, a’-Me2Spm на собственно +1 сдвиг рамки считывания мРНК (Рис. 8). Оказалось,
Рис. 8. Индукция +1 сдвига рамки считывания мРНК сте-реоизомерами MeSpd (1.25 mM) и Me2Spm (2.5 mM). Клетки HEK-293 преинкубировали 48 ч с 2.5 mM DFMO и затем трансфицировали плазмидой, несущей ген люциферазы под контролем промотора, содержащего участок, обеспечивающий +1 сдвиг рамки считывания. Стереоизомеры прибавляли через 12 ч после трансфекции и клетки выращивали еще 24 ч. R, (R)-
a-MeSpd; S, (S)-a-MeSpd; RR, (R,R)-a, a’-Me2Spm; SS, (S,S)- a, a’-Me2Spm; RS, (R,S)- a, a’-Me2Spm.
что наибольшим эффектом среди аналогов Spm обладает (S,S)-a, a’-Me2Spm, а среди анало-гов Spd – (S)-a-MeSpd. Эти данные хорошо объясняют влияние AZ на транспорт аналогов в клетки, а также их эффекты на активность ODC в клетке.
Таким образом, используя оптические изомеры Spm и Spd удалось оригинальным об-разом воздействовать на процессы биосинтеза AZ. Вместе с тем, следует отметить, что моле-кулярные механизмы стереоконтроля +1 сдвига рамки считывания мРНК AZ изомерами Spd и Spm остаются неясными, как и эффекты собственно Spm и Spd, также не имеющие одно-значной интерпретации.
Синтез рацемических b-, g- и w-метилированных аналогов cпермидина,
N1-ацетилспермидина и спермина.
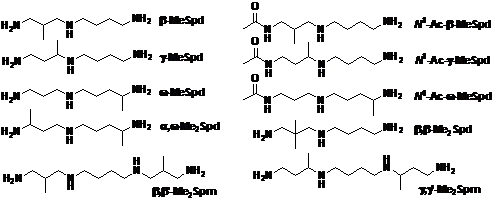 Неизвестные ранее изомеры a-метилированных полиаминов, впервые синтезирован-ные в ходе выполнения настоящего исследования, оказались ценным инструментом исследо-вания клеточных функций Spm и Spd, ферментов их метаболизма и регуляторных механиз-мов, ответственных за поддержание гомеостаза полиаминов в клетке. Оказалось, что конфи-гурация a-углеродного атома “is a hot point of polyamine molecule”. Вместе с тем, вопрос о существовании других критических точек в молекулах Spm и Spd, существенных для кор-ректного узнавания и функционирования полиаминов в клетке, оставался открытым. Более того, учитывая раз-нообразие внутри-клеточных превра-щений Spd и Spm,
Неизвестные ранее изомеры a-метилированных полиаминов, впервые синтезирован-ные в ходе выполнения настоящего исследования, оказались ценным инструментом исследо-вания клеточных функций Spm и Spd, ферментов их метаболизма и регуляторных механиз-мов, ответственных за поддержание гомеостаза полиаминов в клетке. Оказалось, что конфи-гурация a-углеродного атома “is a hot point of polyamine molecule”. Вместе с тем, вопрос о существовании других критических точек в молекулах Spm и Spd, существенных для кор-ректного узнавания и функционирования полиаминов в клетке, оставался открытым. Более того, учитывая раз-нообразие внутри-клеточных превра-щений Spd и Spm,
Рис. 9. Новые С-мети-лированные аналоги Spd, N1-Ac-Spd и Spm.
равно как и множе-ственность их кле-точных функций, представляется вполне обоснованным использовать не только a-метилиро-ванные аналоги, а систему С-метилированных производных (Рис. 9), предполагая, что биохи-миические свойства таких аналогов будут определяться не только трипротонированным при физиологическом рН скелетом Spd, но и положением метильной группы. Эти соображения, несмотря на кажущуюся механистичность перемещения метильной группы по полиаминово-му скелету, позволили получить ряд неожиданных и значимых для биохимии Spm и Spd ре-зультатов (см. следующий раздел).
Для получения b-MeSpd и g-MeSpd удобным представлялось использование восста-новление нитрилов (22а) и (22b), которые были получены присоединением Put к метакрило-нитрилу (21а) и кротононитрилу (21b) (Схема 7). Однако, восстановление нитрилов водоро-дом над Ni-Ренея или PtO2 проходило неоднозначно и сопровождалось образованием набора минорных побочных продуктов, трудно отделяемых от целевых MeSpd’s хроматографией на силикагеле. Тем не менее, этим способом b-MeSpd и g-MeSpd были синтезированы в виде хо-рошо кристаллизующихся трихлоргидратов с суммарными выходами 16% и 32%, считая на метакрилонитрил и кротононитрил, соответственно.
Схема 7

i- NH2(CH2)4NH2/EtOH/20°C→90°C; ii- H2/Ni-Raney/EtOH; iii- Boc2O/THF; iv- LiAlH4/Et2O/-5°C; v- HCl/EtOH.
Для упрощения этих синтезов нитрилы (22а) и (22b) были превращены в соответствующие ди-Вос-производные, которые гладко и с хорошим выходом восстанавливались LiAlH4 в эфире при -5°С до ди-Вос-аминов (23а) и (23b). Последующее удаление защитных групп привело к искомым трихлоргидратам b-MeSpd и g-MeSpd с суммарными выходами 27% и 45%, считая на (21а) и (21b), соответственно.
Промежуточные ди-Вос-амины (23а) и (23b) были использованы в качестве исходных соединений в синтезах N1-ацетильных производных b - и g-MeSpd (Схема 7). Последователь-ное ацетилирование свободной аминогруппы и удаление Вос-защитных групп привели к кристаллическому дихлоргидрату N1-Ас-b-MeSpd с выходом 23.4% и быстро смокающему на воздухе дихлоргидрату N1-Ас-g-MeSpd с выходом 39%, считая на (21а) и (21b).
Учитывая трудности, возникшие при выделении продуктов восстановления нитрилов (22а) и (22b) водородом над Ni-Ренея или PtO2, для получения b, b’-Me2Spm был приготовлен ди-Вос-динитрил (26). Однако этот динитрил не растворялся в эфире, а проведение восста-новления LiAlH4 в THF при -5°С сопровождалось побочным элиминированием. Соответст-венно, после удаления Вос-защитных групп и кристаллизации были получены в примерно равных количествах хорошо кристаллизующийся тетрахлоргидрат b, b’-Me2Spm с суммар-ным выходом 3.5%, считая на метакрилонитрил, и трихлоргидрат b-MeSpd (Схема 8). Для получения g, g’-Me2Spm было использовано восстановление динитрила (24b) водородом над PtO2 (Схема 8), т. к. восстановление ди-Вос-производного динитрила (24b) LiAlH4 в THF при -5°С привело лишь к следовым количествам искомого ди-Вос-диамина. Как и в случае синте-за g-MeSpd (см. выше) наблюдалось образованием спектра побочных продуктов, трудно отде-ляемых от g, g’-Me2Spm хроматографией на силикагеле. Тем не менее, медленно смокающий на воздухе тетрахлоргидрат g, g’-Me2Spm удалось получить с выходом 11.7%, считая на (21а).
Схема 8
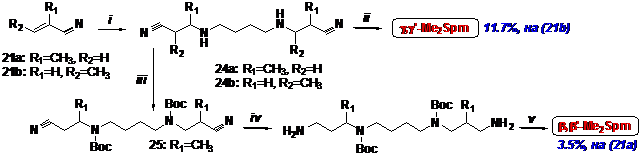
i- NH2(CH2)4NH2/20°C→90°C; ii- H2/PtO2/EtOH; iii- Boc2O/THF; iv- LiAlH4/Et2O/-5°C; v- HCl/EtOH.
w-MeSpd и a, w-Me2Spd сначала были синтезированы исходя из 3-аминобутанола-1 (2) (Схема 9). Ключевым интермедиатом этих 12-ти стадийных синтезов был Вос-нозилат (28), который алкилировали 3-(N-бензилоксикарбонил)амино-1-бромпропаном для получе-ния (29а), являющегося предшественником w-MeSpd, или 3-(N-бензилоксикарбонил)амино-1-бромбутаном для получения (29b) – предшественника a, w-Me2Spd (Схема 9). Последую-
Схема 9

i- Boc2O/THF; ii- MsCl/Et3N/CH2Cl2; iii- NaCN/DMSO; iv- LiAlH4/Et2O/-5°C; v- NsCl/CH2Cl2/Et3N;
vi- 1. CbzNH(CH2)3Br/DMF/K2CO3/45°C для 28а; 2. CbzNHCH(CH3)(CH2)2Br/DMF/K2CO3/45°C для 28b;
vii- PhSH/DMF/K2CO3; viii- 1. HCl/EtOH; 2. H2/Pd/AcOH/MeOH;HCl/EtOH.
щее удаление защитных групп гладко приводило к целевым хорошо кристаллизующимся трихлоргидратам w-MeSpd и a, w-Me2Spd с суммарными выходами 31%, считая на метан-сульфонат (27) и 12.2%, считая на аминоспирт (2), соответственно.
Впоследствии (Схема 10), для упрощения синтеза w-MeSpd мы использовали алкили-
Схема 10
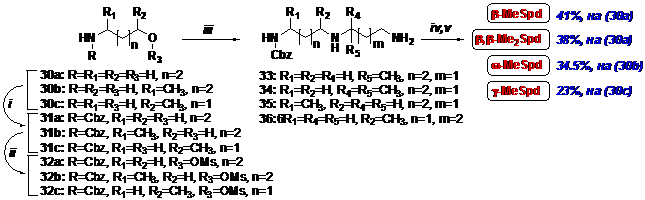
i- CbzCl/H2O/NaHCO3; ii- MsCl/Et3N/ CH2Cl2; iii- 1. NH2CH2CH(CH3)CH2NH2 для b-MeSpd;
2. NH2CH2C(CH3)2CH2NH2 для b, b-Me2Spd; 3. NH2(CH2)3NH2/THF для w-MeSpd; 4. NH2(CH2)4NH2/THF
для g-MeSpd; iv- H2/Pd/MeOH/AcOH; i- HCl/EtOH.
рование диаминов мезилатами N-защищенных спиртов. Этим методом в 4 стадии были по-лучены также b-MeSpd, b, b’-MeSpd и g-MeSpd с суммарными выходами 41%, 38% и 23%, считая на (30а) и (30с), соответственно. Выход w-MeSpd составил 34.5%, считая на (30b).
Промежуточный Cbz-диамин (35) был использован в качестве исходного соединения при получении N1-Ас-w-MeSpd (Схема 11). Для дискриминации первичной и вторичной ами-ногрупп было использовано превращение первичной в салицилиденовое производное, кото-рое после введения Вос-защиты по вторичной аминогруппе, было превращено в первичный амин (36) обработкой CH3ONH2-основанием. Ацетилирование первичной аминогруппы и последовательное удаление Boc - и Cbz-защитных групп привели к целевому N1-Ac-w-MeSpd с выходом 46%, считая на (35).
Схема 11
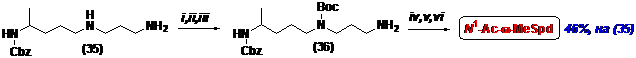
i- o-C6H4(OH)CHO/THF; ii- Boc2O/THF; iii- MeONH2/THF; iv- AcCl/Et3N/THF; v- H2/Pd/AcOH/MeOH; vi- HCl/EtOH
Таким образом, семейство a-метилированных аналогов Spm и Spd было дополнено неизвестными ранее b-, g-, w - и a, w-Me2Spd, N1-ацетильными производными b-, g- и w-MeSpd, а также b, b’-Me2Spm и g, g’-Me2Spm.
Взаимодействие С-метилированных аналогов Spd с ферментами
метаболизма полиаминов и клетками [‡]
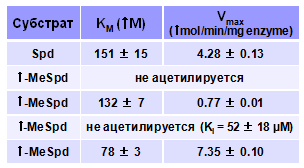 На первом этапе мы исследовали взаимодействие b-MeSpd, g-MeSpd, w-MeSpd и a, w-Me2Spd со SSAT в сравнении с a-MeSpd и Spd. Как и следовало ожидать, w-MeSpd, подобно описанному ранее w, w-Me2Spd (Nagarajan S., et al. 1988. Biochem.J., 254, 373-378), оказался хорошим субстратом фермента, а a, w-Me2Spd не ацетилировался как и a-MeSpd (Табл. 5). Удивительно, но b-MeSpd, в отличие от описанного ранее b, b-Me2Spd, имеющего две метиль-ные группы при b-углеродном атоме и a-MeSpd, оказался хорошим субстратом SSAT с близким Spd значением Km, но заметно снижен-ным, по сравнению со Spd значением Vmax.
На первом этапе мы исследовали взаимодействие b-MeSpd, g-MeSpd, w-MeSpd и a, w-Me2Spd со SSAT в сравнении с a-MeSpd и Spd. Как и следовало ожидать, w-MeSpd, подобно описанному ранее w, w-Me2Spd (Nagarajan S., et al. 1988. Biochem.J., 254, 373-378), оказался хорошим субстратом фермента, а a, w-Me2Spd не ацетилировался как и a-MeSpd (Табл. 5). Удивительно, но b-MeSpd, в отличие от описанного ранее b, b-Me2Spd, имеющего две метиль-ные группы при b-углеродном атоме и a-MeSpd, оказался хорошим субстратом SSAT с близким Spd значением Km, но заметно снижен-ным, по сравнению со Spd значением Vmax.
Табл. 5. Взаимодействие C-метилированных анало-гов Spd с SSAT.
Таким образом, уменьшение количества замес-тителей кардинально влияет на субстратные свойства аналога. Оказалось, что перемещение метильной группы еще на одно положение, приводящее к g-MeSpd, вновь лишает аналог суб-стратных свойств в SSAT-реакции (Табл. 5). Такие, на первый взгляд, парадоксальные зави-симости субстратных свойств от положения и количества метильных групп хорошо согласу-ются с данными рентгено-структурных исследований комплекса фермента с коньюгатом Spm и СоА, моделирующим бисубстратный комплекс (Hegde S. S., et al. 2007. Biochemistry, 46, ), а также бинарным комплексом SH-CoA – Spm (Montemayor E. J. & Hoffman D. W. 2008. Biochemistry, 47, 9145–9153) и отражают особенности узнавания полиаминов SSAT.
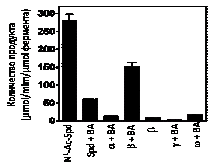 В предыдущем разделе было показано, что N-Ac-a-MeSpd является хорошим субстра-том АРАО, а в присутствии ароматических альдегидов АРАО эффективно расщепляла и соб-ственно a-MeSpd. Поэтому мы исследовали возможность окислительного расщепления С-метилированных MeSpd’s под действием АРАО в присутствии бензальдегида, который по данным (Holtta E. 1977. Biochemistry, 16, 91–100) является самым активным из исследован-ных альдегидов. Оказалось, что бензальдегид эффективно стимулирует катализируемое АРАО расщепление всех синтезированных аналогов за исключением g-MeSpd, в котором метильная группа находится при атоме углерода, образующего расщепляемую в ходе поли-аминооксидазной реакции связь со вторичной аминогруппой (Рис. 10). Соответственно, этот участок субстрата должен быть высококомплементарен активному центру фермента и нали-чие метильной группы в g-положении Spd может привести к непродуктивному связыванию этого аналога или затруднить отрыв протона, являющийся движущей силой окислительного расщепления, приводящего к образованию промежуточного основания Шиффа. Таким образом, g-MeSpd не является субстратом ни АРАО, ни SSAT и представляет собой новый катаболически устойчивый аналог Spd.
В предыдущем разделе было показано, что N-Ac-a-MeSpd является хорошим субстра-том АРАО, а в присутствии ароматических альдегидов АРАО эффективно расщепляла и соб-ственно a-MeSpd. Поэтому мы исследовали возможность окислительного расщепления С-метилированных MeSpd’s под действием АРАО в присутствии бензальдегида, который по данным (Holtta E. 1977. Biochemistry, 16, 91–100) является самым активным из исследован-ных альдегидов. Оказалось, что бензальдегид эффективно стимулирует катализируемое АРАО расщепление всех синтезированных аналогов за исключением g-MeSpd, в котором метильная группа находится при атоме углерода, образующего расщепляемую в ходе поли-аминооксидазной реакции связь со вторичной аминогруппой (Рис. 10). Соответственно, этот участок субстрата должен быть высококомплементарен активному центру фермента и нали-чие метильной группы в g-положении Spd может привести к непродуктивному связыванию этого аналога или затруднить отрыв протона, являющийся движущей силой окислительного расщепления, приводящего к образованию промежуточного основания Шиффа. Таким образом, g-MeSpd не является субстратом ни АРАО, ни SSAT и представляет собой новый катаболически устойчивый аналог Spd.
Рис. 10. Взаимодействие C-метилированных аналогов Spd с APAO в присутствии или без 5 mM бензальдегида (BА).
Следует особо отметить, что b-MeSpd достаточно эффектив-но расщеплялся АРАО и в отсутствии бензальдегида (Рис. 10), что совершенно не характерно для этого фермента, учитывая известные данные о его субстратной специфичности (см. обзор: Casero R. A. & Pegg A. E. 2009. Biochem.J., 421, 323–338). В присутствии же бензальдегида, субстратные свойства b-MeSpd намного превосходили таковые для Spd в аналогичных усло-виях (Рис. 10). Таким образом, перемещая метильную группу по скелету спермидина, удается получать как катаболически устойчивые производные (g-MeSpd), так и усиливать способ-ность аналога к окислительному расщеплению (b-MeSpd), то есть появляются неизвестные ранее возможности для регуляции метаболизма полиаминов.
На следующем этапе был исследован транспорт синтезированных аналогов Spd в клетки DU145. Все производные узнавались системой транспорта в качестве Spd (Рис. 11) и конкурировали за транспорт с [14C]-Spd, однако лишь b-MeSpd был столь же эффективен как
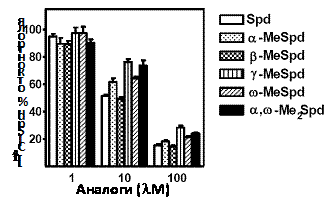 и “холодный“ Spd, тогда как остальные С-метилированные производные оказались несколь-ко менее активными.
и “холодный“ Spd, тогда как остальные С-метилированные производные оказались несколь-ко менее активными.
Рис. 11. Транспорт C-метилированных аналогов Spd в клетки DU145. Клетки инкубировали в течение 10 мин с 10 mM [14C]-Spd в присутствии 1, 10 или 100 mM исследуемых аналогов.
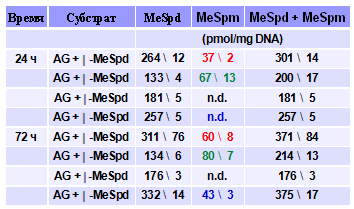 Проникнув в клетку С-метилирован-ные производные Spd могут превращаться в соответствующие аналоги Spm. Из литера-туры известно, что гем-Me2Spd’s, несущие две метильные группы при одном атоме углерода (Nagarajan S., et al. 1988. Biochem.J., 254, 373-378), не являются субстратами Spm-синтазы. Уменьшение количества метильных групп, как и в случае SSAT (см. выше), кардинально сказывается на субстратных свойствах аналогов В клетках DU145 a-MeSpd, b-MeSpd и w-MeSpd метаболизировали до соответствующих про-изводных Spm (Табл. 6), но этого не наблюдалось в клетках дефицитных по Spm-синтазе (данные не приведены). Следует отметить, что a-MeSpm и b-MeSpm образовывались уже после 24 ч инкубации клеток с соответ-ствующими аналогами Spd, тогда как d-MeSpm, возникающий из w-MeSpd де-тектировался лишь после 48 ч инкуба-ции аналога с клетками.
Проникнув в клетку С-метилирован-ные производные Spd могут превращаться в соответствующие аналоги Spm. Из литера-туры известно, что гем-Me2Spd’s, несущие две метильные группы при одном атоме углерода (Nagarajan S., et al. 1988. Biochem.J., 254, 373-378), не являются субстратами Spm-синтазы. Уменьшение количества метильных групп, как и в случае SSAT (см. выше), кардинально сказывается на субстратных свойствах аналогов В клетках DU145 a-MeSpd, b-MeSpd и w-MeSpd метаболизировали до соответствующих про-изводных Spm (Табл. 6), но этого не наблюдалось в клетках дефицитных по Spm-синтазе (данные не приведены). Следует отметить, что a-MeSpm и b-MeSpm образовывались уже после 24 ч инкубации клеток с соответ-ствующими аналогами Spd, тогда как d-MeSpm, возникающий из w-MeSpd де-тектировался лишь после 48 ч инкуба-ции аналога с клетками.
Табл. 6. Биосинтез C-метилированных анало-гов Spm в клетках DU145. Клетки выращивали в присутствии C-метилированных аналогов Spd (100 mM) и 1 mM аминогуанидина (AG), ингиби-тора сывороточных аминооксидаз, предотвраща-ющего неспецифическую токсичность аналогов.
Удивительно, что g-MeSpd не метаболизировал в клетках DU145 до g-MeSpm даже после 72 ч инкубации клеток с аналогом (Табл.6), что прямо указывает на существование высокой комплементарности между соответствующим участком активного центра Spm-синтазы и центральным фрагментом молекулы Spd, обеспечивающим продуктивное связывание суб-страта и его аналогов в активном центре фермента. Таким образом, g-MeSpd представляет собой новый метаболически устойчивый аналог Spd, а перемещение метильной группы по скелету Spd позволяет регулировать субстратные свойства С-метилированных производных Spd в Spm-синтазной реакции, что не описано ни для одного из известных аналогов Spd.
Сбалансированная работа ферментов синтеза и деградации полиаминов наряду с сис-темой активного транспорта Spm и Spd, являются основными факторами, позволяющими поддерживать необходимую концентрацию полиаминов в клетке. Активность, биосинтез и деградация ферментов метаболизма полиаминов зависят от многих факторов, в том числе и
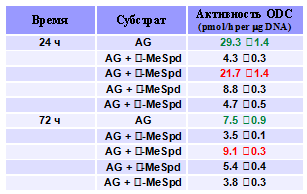 от внутриклеточного содержания Spm и Spd. Поэтому было изучено влияние С-метилирован-ных аналогов Spd на активность ключевых ферментов метаболизма полиаминов. Наиболее выраженные эффекты наблюдались в случае ODC – скорость-лимитирующего фермента на путях биосинтеза полиаминов (Табл. 7). Оказа-лось, что b-MeSpd, в противоположность всем остальным производным, практически не сни-жал активность ODC ни после 24 ч, ни после 72 ч инкубации клеток с аналогом.
от внутриклеточного содержания Spm и Spd. Поэтому было изучено влияние С-метилирован-ных аналогов Spd на активность ключевых ферментов метаболизма полиаминов. Наиболее выраженные эффекты наблюдались в случае ODC – скорость-лимитирующего фермента на путях биосинтеза полиаминов (Табл. 7). Оказа-лось, что b-MeSpd, в противоположность всем остальным производным, практически не сни-жал активность ODC ни после 24 ч, ни после 72 ч инкубации клеток с аналогом.
Табл.7. Влияние C-метилированных аналогов Spd (100 µM) на активность ODC в клетках DU145. Клетки выра-щивали в присутствии 1 mM AG.
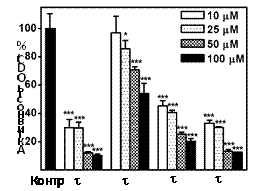 Такие необычные свойства b-MeSpd привели к необходимости оценить концентрационную и временную зависимости эффекта. Оказалось, что в случае 4-х ч инкубации клеток DU145 с 10 μM b-MeSpd активность ODC практически не изменялась, тогда как для a-MeSpd она сос-тавляла ~30%, для w-MeSpd ~40%, а для g-MeSpd – около 50% от активности ODC в конт-рольных клетках (Рис. 12). Подобная зависимость активности аналога от положения метиль-ной группы является уникальной и связана, скорее все-го, с низкой эффективностью индукции биосинтеза AZ b-MeSpd, что совершенно не характерно для функцио-нально-активных миметиков полиаминов, и открывает спектр новых возможностей для регулирования мета-болизма полиаминов.
Такие необычные свойства b-MeSpd привели к необходимости оценить концентрационную и временную зависимости эффекта. Оказалось, что в случае 4-х ч инкубации клеток DU145 с 10 μM b-MeSpd активность ODC практически не изменялась, тогда как для a-MeSpd она сос-тавляла ~30%, для w-MeSpd ~40%, а для g-MeSpd – около 50% от активности ODC в конт-рольных клетках (Рис. 12). Подобная зависимость активности аналога от положения метиль-ной группы является уникальной и связана, скорее все-го, с низкой эффективностью индукции биосинтеза AZ b-MeSpd, что совершенно не характерно для функцио-нально-активных миметиков полиаминов, и открывает спектр новых возможностей для регулирования мета-болизма полиаминов.
Рис. 12. Влияние C-метилированных аналогов Spd на актив-ность ODC в клетках DU145. Клетки инкубировали с 10, 25, 50 или 100 mM аналогов и 1 mM AG в течение 4 ч.
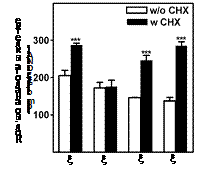 На следующем этапе был исследован транспорт С-метилированных аналогов Spd в об-работанные и необработанные СНХ (ингибитор биосинтеза белка) клетки DU145. Оказалось, что в отсутствии AZ все аналоги за исключением b-MeSpd накапливались значительно эф-фективнее, чем в клетках (Рис. 13), способных синтезировать дополнительные количества AZ в ответ на повышение внутриклеточной концентрации аналогов полиаминов, что пре-пятствует проникновению экзогенных полиаминов в клетки и способствует поддержанию гомеостаза Spm и Spd. В случае b-MeSpd различий между обработанными и необработанными
На следующем этапе был исследован транспорт С-метилированных аналогов Spd в об-работанные и необработанные СНХ (ингибитор биосинтеза белка) клетки DU145. Оказалось, что в отсутствии AZ все аналоги за исключением b-MeSpd накапливались значительно эф-фективнее, чем в клетках (Рис. 13), способных синтезировать дополнительные количества AZ в ответ на повышение внутриклеточной концентрации аналогов полиаминов, что пре-пятствует проникновению экзогенных полиаминов в клетки и способствует поддержанию гомеостаза Spm и Spd. В случае b-MeSpd различий между обработанными и необработанными
Рис. 13. Влияние AZ на транспорт C-метилированных аналогов Spd в клетки DU145. Клетки инкубировали 2 ч с CHX (10 mg/mL), или без CHX, а затем еще 4 ч с аналогами Spd (100 mM).
СНХ клетками не наблюдалось (Рис. 13), что косвенно указы-зывает на то, что этот аналог, в отличие от других С-метилированных производных Spd не
способен, или весьма слабо индуцирует биосинтез AZ в клетках DU145.
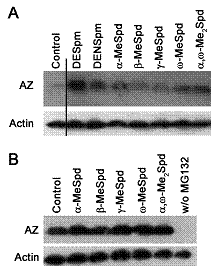 Способность С-метилированных производных Spd индуцировать биосинтез AZ по-добно природным полиаминам и большинству их синтетических аналогов, была оценена ис-пользуя антитела к AZ. Оказалось, что количество AZ зави-сит от положения метильной группы в структуре аналога. b-MeSpd слабо индуцировал биосинтез AZ, g-MeSpd был нес-колько активнее, но действовал слабее чем остальные С-ме-тилированные производные Spd и контрольные DENSpm и DESpm (Рис. 14A). Пониженная способность b-MeSpd инду-
Способность С-метилированных производных Spd индуцировать биосинтез AZ по-добно природным полиаминам и большинству их синтетических аналогов, была оценена ис-пользуя антитела к AZ. Оказалось, что количество AZ зави-сит от положения метильной группы в структуре аналога. b-MeSpd слабо индуцировал биосинтез AZ, g-MeSpd был нес-колько активнее, но действовал слабее чем остальные С-ме-тилированные производные Spd и контрольные DENSpm и DESpm (Рис. 14A). Пониженная способность b-MeSpd инду-
Рис. 14. Влияние C-метилированных аналогов Spd на биосинтез AZ. Клетки инкубировали 4 ч с исследуемыми аналогами (100 mM) без MG132 (A) или в присутствии 25 mM ингибитора (B). Аликвоту суммар-ной белковой фракции (50 mg) анализировали электрофорезом в 12 % SDS-полиакриламидном геле и визуализировали иммуноблотингом с антителами к AZ.
цировать биосинтез AZ была подтверждена и в экспериментах с MG132, ингибитором про-теосом-опосредованной деградации белков. В присутствии MG132 количество AZ сущест-венно увеличивалось после добавления в среду любого из С-метилированных производных MeSpd, за исключением b-MeSpd (Рис 14В). Возможность регулировать экспрессию AZ простым перемещением метильной группы вдоль скелета Spd является совершенно неожи-данной, уникальной и не имеющей аналогий в литературе, так как все исследованные функ-ционально-активные аналоги Spm и Spd вызывают +1 сдвиг рамки считывания мРНК AZ и индуцируют его биосинтез (см. обзор: Kahana С. 2009. Cell.Mol.Life Sci., 66, ).
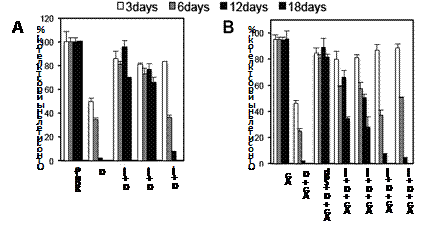 На заключительном этапе была исследована способность С-метилированных аналогов Spd восстанавливать рост клеток DU145 с истощенным пулом полиаминов (18 сут инкубации с 5 mM DFMO) (Рис. 15). Из литературы известно, что даже гем-Me2Spd’s способны поддер-живать рост клеток с острым дефицитом полиаминов, возникающим в результате инкубации с DFMO в течение 3 сут, т. е. в условиях когда еще не затра-гиваются процессы гипузини-лирования eIF5А
На заключительном этапе была исследована способность С-метилированных аналогов Spd восстанавливать рост клеток DU145 с истощенным пулом полиаминов (18 сут инкубации с 5 mM DFMO) (Рис. 15). Из литературы известно, что даже гем-Me2Spd’s способны поддер-живать рост клеток с острым дефицитом полиаминов, возникающим в результате инкубации с DFMO в течение 3 сут, т. е. в условиях когда еще не затра-гиваются процессы гипузини-лирования eIF5А

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
