
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
НОВЫЕ ПОДХОДЫ К ХИМИЧЕСКОМУ РЕГУЛИРОВАНИЮ МЕТАБОЛИЗМА БИОГЕННЫХ АМИНОВ СПЕРМИНА И СПЕРМИДИНА
03.01.03 - Молекулярная биология
Автореферат
диссертации на соискание ученой степени
доктора химических наук
Москва 2011
Работа выполнена в Лаборатории молекулярных основ действия физиологически
активных соединений Учреждения Российской академии наук Института молекулярной биологии им. РАН
Официальные оппоненты: д. х.н., проф., чл.-корр. РАН
д. х.н., проф., чл.-корр. РАН
д. х.н., проф.
Ведущая организация: Учреждения Российской академии наук
Институт биохимии им. А.Н. Баха РАН
Защита диссертации состоится « 24 » ноября 2011 г. в 11-00 часов на заседании диссертационного Совета Д 002.235.01 при Учреждении Российской академии наук Институте молекулярной биологии им. РАН Москва, ул. Вавилова, д. 32.
С диссертацией можно ознакомиться в библиотеке Учреждения Российской академии наук Институте молекулярной биологии им. РАН
Автореферат разослан « ____ » _____________ 2011 г.
Ученый секретарь диссертационного Совета
кандидат химических наук
Актуальность проблемы. Биогенные полиамины спермин (Spm), спермидин (Spd) и их предшественник путресцин (Put) присутствуют в клетках всех типов в mM-mM концентра-циях и жизненно необходимы для их нормального роста. Высокое внутриклеточное содержа-ние Spm и Spd определяет разнообразие их клеточных функций, многие из которых остаются все еще малоизученными, что обеспечивает поступательное развитие этой области биохимии (см. обзор: Casero R. A. & Pegg A. E. In: Polyamines: Methods and Protocols, Methods in Molecu-lar Biology, 720, 3-35. Springer Science+Business Media, LLC. 2011). При физиологическом значении рН полиамины существуют в форме поликатионов. Spd и Spm взаимодействуют с ДНК, РНК и нуклеопротеидами, служат регуляторами активности топоизомераз, рестриктаз, а также ферментов биосинтеза ДНК и РНК. Полиамины участвуют в регуляции транспорта Са2+ митохондриями, являются модуляторами NMDA рецептора и эффекторами транспорта K+, а также регулируют процесс полимеризации тубулина. Spd служит единственным доно-ром аминобутильного остатка, необходимого для посттрансляционной модификации (прев-ращение остатка лизина в гипузин – Ne-(4-амино-2-оксибутил)-L-лизин) фактора инициации трансляции eIF-5A, жизненно необходимого для всех эукариот. Наконец, Spd входит в состав трипанотиона – важнейшего метаболита протозойных паразитов.
Аналоги Spm и Spd являются одним из основных инструментов исследования фермен-тов метаболизма и клеточных функций полиаминов. Соответственно, направленное создание новых биологически активных производных полиаминов, расширение семейства весьма не-многочисленных функционально-активных миметиков частично взаимозаменяемых Spm и Spd, изучение взаимодействия этих аналогов с ферментами метаболизма полиаминов, а так-же их активности на уровне культуры клеток и in vivo представляется вполне актуальным.
Целью работы являлись: (i)- разработка удобных способов получения системы неиз-вестных ранее С-метилированных аналогов Spd, N1-ацетил-Spd и Spm; (ii)- синтез неописан-ных ранее оптических изомеров a-метилполиаминов; (iii)- создание нового типа зарядодефи-цитного аналога Spm (SpmTrien) на основе триэтилентетрамина, а также ацетильных и бис-этильных производных SpmTrien; (iv)- получение оригинальных биологически активных гид-роксиламин содержащих аналогов Spm и агматина (последнее потребовало создания нового метода гуанидилирования аминооксигруппы); (v)- исследование взаимодействия С-метили-рованных аналогов полиаминов и SpmTrien с ДНК; (vi)- изучение взаимодействия синтезиро-ванных веществ с ферментами метаболизма полиаминов и их эффектов в культуре клеток и in vivo.
Научная новизна. Разработаны удобные схемы синтеза системы неизвестных ранее С-метилированных аналогов Spd, Spm и N1-ацетил-Spd, позволяющие получать эти соедине-ния с высокими выходами из доступных исходных веществ и в количествах, необходимых для проведения экспериментов in vivo. Получены неизвестные ранее оптические изомеры a-метилполиаминов и N-ацетил-a-MeSpd. Синтезирован оригинальный зарядодефицитный аналог Spm – SpmTrien и его биохимически-значимые производные. Разработаны удобные схемы синтеза аминоокси - и окса-аналогов Spm, а также бисубстратных ингибиторов спермин/спермидин-N1-ацетилтрансферазы (SSAT) на основе аминоокси аналогов Spd и Spm. Впервые показано, что трифлат ди-Вос-гуанидина количественно гуанидилирует аминооксигруппу, что позволило получить ряд функционально-замещенных алкоксигуани-динов, включая и неизвестные ранее гидроксиламин-содержащие аналоги агматина.
Исследовано взаимодействие синтезированных С-метилированных производных Spd и Spm с ферментами метаболизма полиаминов, их влияние на антизим-зависимые пути регу-ляции гомеостаза полиаминов, а также способность аналогов восстанавливать рост клеток с истощенным пулом полиаминов. Оказалось, что биохимические свойства аналогов определя-ются положением метильной группы. Введение метильной группы в a-положение обеспечи-вает катаболическую устойчивость a-метилспермидина (a-MeSpd), но не a-метилспермина (a-MeSpm). Показано, что среди аналогов этого типа лишь g-метилспермидин (g-MeSpd) является функционально-активным метаболически устойчивым миметиком спермидина (g-MeSpd не является субстратом ни SSAT, ни сперминсинтазы, но способен восстанавливать рост клеток с истощенным пулом полиаминов), что открывает новые возможности для иссле-дования клеточных функций катаболически неустойчивых и легко взаимопревращающихся Spm и Spd. Найдено, что b-метилспермидин (b-MeSpd), в отличие от подавляющего боль-шинства аналогов Spd и остальных С-метилированных производных Spd, практически не ин-дуцирует биосинтез антизима – небольшого белка блокирующего проникновение полиами-нов в клетку, а также вызывающего диссоциацию орнитиндекарбоксилазы на субъединицы и осуществляющего их доставку в 26S протеосомы. Использование хиральных аналогов a-ме-тилированных полиаминов позволило впервые показать, что сперминоксидаза, дезоксигипу-зинсинтаза и ацетилполиаминооксидаза (АРАО), природные субстраты которых ахиральны, обладают стереоспецифичностью и найти изящный способ регулирования стереоспецифич-ности АРАО. Установлено, что a-метилполиамины способны выполнять жизненно важные функции их природных прототипов в культуре нормальных и опухолевых клеток, а также и in vivo, предотвращая развитие острого панкреатита у SSAT-трансгенных крыс и стимулируя регенерацию печени после частичной гепатэктомии в условиях дефицита Spm и Spd. Таким образом, впервые показано, что введение метильной группы в заданное положение поли-аминного скелета и изменение конфигурации хирального центра позволяет тонко регулиро-вать биохимические свойства аналогов полиаминов, а соответствующие производные и ана-логи представляют собой ценные инструменты исследования клеточных функций и метабо-лизма Spm и Spd in vitro и in vivo.
Практическая ценность работы. Повышенное содержание Spm и Spd в опухолевых клетках, жизненная необходимость полиаминов для развития возбудителей малярии, сонной болезни и лейшманиоза, а также наблюдаемые нарушения метаболизма полиаминов у боль-ных панкреатитом и диабетом формируют прикладную составляющую исследований клеточ-ных функций Spm и Spd и поиска новых регуляторов их метаболизма. Правомерность подоб-ного подхода подтверждается успешным использованием a, a-дифторметилорнитина (DFMO) для лечения поздних стадий сонной болезни (Burri C. C. & Brun R. 2003. Parasitol. Res. 90, S49-S52) и противоопухолевой активностью терминально N,N’-бис-алкилированных производных полиаминов, являющихся супериндукторами SSAT (см. обзор: Casero R. A. & Marton L. J. 2007. Nat. Rev. Drug Discov. 6, 373-390). В настоящей работе впервые показано, что a-MeSpd спосо-бен предотвращать развитие острого панкреатита у SSAT-трансгенных крыс, возникающего в ответ на активацию фермента. Найдено, что активно транспортируемый в L.donovani 1-гуани-диноокси-3-аминопропан (GAPA), в отличие от других ингибиторов орнитиндекарбоксилазы, эффективно препятствует размножению резистентных к сурьма-содержащим препаратам форм паразита, что открывает новые возможности для практической полиаминотерапии лейшманиоза.
Апробация работы. Отдельные части работы докладывались на: Polyamine Gordon Research Conference (Connecticut College, New London, CT, USA, June 2001); International Con-ference devoted to 100-year anniversary of Acad. A. E.Braunstein (Moscow, Russia, May 2002); International Conference on Biogenic Amines (Trento, Italy, June 2002); Biogenic Amines COST 922: Health implications of dietary amines (Trento, Italy, May 2004); The IX spring meeting of synthetic chemistry (Kuopio, Finland, June 2006); 1st International Conference on the Role of Poly-amines and their Analogs in Cancer and Other Diseases (Tivoli, Italy, September 2006); Biochemi-cal Society COST Action 922: Health Implications of Dietary Amines (Aberdeen, Scotland, UK, October 2006); COST B22 expert meeting "Polyamines and parasites" (Stockholm, Sweden, Sep-tember 2007); International Conference “Polyamine: Forty Years of Mammalian Ornithine Decar-boxylase” (Kuopio, Finland, June 2008); XI International Congress on Amino Acids, Peptides and Proteins (Vienna, Austria, August 2009); 2nd International Conference on the Role of Polyamines and their Analogs in Cancer and other Diseases (Tivoli, Italy, December 2010); Polyamine Gordon Research Conference (Waterville Valley, New Hampshire, USA, June 2011); International Conference “Biogenic Amines 2011. Biochemical, Physiological and Clinical Perspectives” (Trento, Italy, September 2011).
Публикации. По материалам диссертации опубликовано 37 статей.
Структура и объем диссертации. Диссертация изложена на ___ стр. машинописного текста и состоит из введения, обзора литературы, обсуждения полученных результатов, экс-периментальной части и выводов. Материал иллюстрирован ___ таблицами, ___ рисунками и ___ схемами. Список цитированной литературы включает ___ наименований.
Работа выполнена при финансовой поддержке грантами РФФИ: , , -ИНД, -АФ, , -Инд; а также Федераль-ной целевой программой “Научные и научно-педагогические кадры инновационной России” (Госконтракт № 02.740.11.5134 от 09 марта 2010 г.) и Программой президиума РАН "Моле-кулярная и клеточная биология"
Система метаболизма полиаминов включает в себя несколько ферментов (Рис. 1), ско-рость определяющими из которых на путях биосинтеза являются декарбоксилазы орнитина (ODC) и S-аденозилметионина (AdoMetDC), а на путях катаболизма – спермидин/спермин-N1-ацетилтрансфераза (SSAT). Биосинтез и деградация этих трех короткоживущих фермен-тов тонко регулируется на многих уровнях, в том числе и в ответ на изменения внутрикле-точной концентрации полиаминов. Повышение содержания спермина (Spm) и спермидина (Spd) вызывает +1 сдвиг рамки считывания мРНК антизима (AZ), что приводит к образова-нию функционального активного белка, который блокирует проникновение полиаминов в клетку, а также, связываясь с ODC, вызывает диссоциацию фермента на субъединицы и осу-ществляет их доставку в 26S протеосомы. Повышение концентрации Spm и Spd вызывает продуктивный сплайсинг мРНК SSAT, что, вместе с AZ-зависимыми механизмами регуля-ции, является одним из основных способов поддержания гомеостаза полиаминов в клетке.
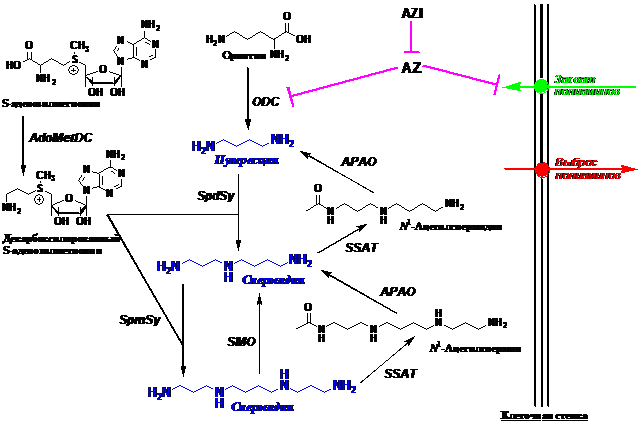
Рис.1. Метаболизм полиаминов. ODC – орнитиндекарбоксилаза; AdoMetDC – S-аденозилметиониндекарбок- силаза; SSAT – спермидин/спермин-N1-ацетилтрансфераза; АPAО – ацетилполиаминоксидаза; SMO – спермин-оксидаза; SpdSy – спермидинсинтаза; SpmSy – сперминсинтаза; AZ – антизим; AZI – ингибитор антизима.
Учитывая, что опухолевые клетки имеют повышенный уровень полиаминов по срав-нению с нормальными, а также необходимость полиаминов для размножения болезнетвор-ных трипаносоматидов (Plasmodium falciparum, Trypanosoma brucei gambiense, Leishmania donovani и т. п.) и то, что Spm и Spd входят в состав оболочки многих вирусов, в биохимии полиаминов большое внимание традиционно уделяется разработке подходов к истощению внутриклеточного пула Spd и Spm, приводящего к ингибированию роста клеток. Соответст-венно были созданы разнообразные специфические ингибиторы ферментов биосинтеза по-лиаминов (см. обзоры: Wallace H. M. & Fraser A. V. 2004. Amino Acids, 26, 353–365; Seiler N. 2003. Curr.Drug Targets, 4, 537-564). В культуре клеток экзогенные Spm и Spd полностью об-ращают эффекты ингибиторов ферментов биосинтеза полиаминов, что подтверждает избира-тельность исходного воздействия на систему. Однако наиболее эффективным способом сни-жения уровня полиаминов in vitro и in vivo оказалась супериндукция SSAT – скорость опре-деляющего фермента катаболизма полиаминов (см. обзоры: Casero R. A. & Woster P. M. 2009. J. Med. Chem., 52, ; Casero R. A. & Marton L. J. 2007. Nat. Rev. Drug Discov. 6, 373-390). Так как в условиях супериндукции SSAT экзогенные Spm и Spd быстро катаболизируют и поэтому неэффективны, то оставалось неясным, обусловлены ли наблюдаемые эффекты только снижением уровней Spm/Spd в результате супериндукции SSAT или же они прямо не связаны с изменением содержания полиаминов в соответствующих органах и тканях. Для от-вета на этот важнейший для биохимии полиаминов вопрос необходимо было создать систему катаболически устойчивых аналогов Spm и Spd, способных выполнять основные клеточные функции полиаминов. В общем смысле, значение подобных веществ велико и потому, что регуляция биохимических процессов подразумевает не только возможности вызывать необ-ходимый ответ клетки или организма, но и наличие в распоряжении исследователя способов обращения эффекта. Это позволяет оценить избирательность исходного воздействия и свиде-тельствует об адекватности наших представлений о том или ином биохимическом процессе.
Изучение клеточных функций полиаминов на молекулярном уровне осложнено час-тичной взаимозаменяемостью и легкостью взаимопревращений Spm и Spd. Поэтому в этих исследованиях широко используются мутантные микроорганизмы и клетки (Cohen S. S. A guide to the polyamines. New York: Oxford University Press. 1998), а также трансгенные живот-ные (см. обзор: Janne J., et al. 2004. Eur.J.Biochem., 271, 877-894), что позволяет активировать или «выключать» заданные ферменты метаболизма полиаминов и регуляторные механизмы, ответственные за поддержание гомеостаза полиаминов. Относительно простым решением, хорошо дополняющим существующие подходы и позволяющим дискриминировать клеточ-ные эффекты Spm и Spd, может быть истощение внутриклеточного пула полиаминов и ис-пользование для обращения эффекта функционально активных миметиков Spm и Spd не спо-собных к взаимопревращениям. Однако соединения с подобным комплексом свойств, равно как и функционально-активные аналоги Spm и Spd не индуцирующие биосинтез AZ, до настоящего времени не были известны.
Совокупность этих обстоятельств привела к синтезу системы неизвестных ранее С-ме-тилированных аналогов Spm и Spd, включая оптические изомеры a-метилированных произ-водных, и исследованию их взаимодействия с ферментами метаболизма полиаминов, а также эффектов в культуре клеток и in vivo, предполагая, что конкретные биохимические свойства аналогов будут определяться не только протонированным при физиологическом рН скелетом молекулы полиамина, но и положением метильной группы, а также и конфигурацией возни-кающего хирального центра. Результаты этих исследований представлены в первой части диссертационной работы.
Биохимические свойства Spm и Spd в значительной степени определяются правильным расположением аминогрупп, протонированных при физиологическом значении рН. Однако вклад зарядовой составляющей, по сравнению со структурным фактором, в активность полиаминов малоисследован. Рациональным подходом для изучения вклада протонирования аминогрупп в функционирование полиаминов является констуирование изостерных “зарядодефицитных” аналогов – рКа одной или двух аминогрупп в таких производных понижена на несколько порядков по сравнению с аминогруппами Spm и Spd. Соответственно, во второй части настоящей работы предложен и реализован оригинальный принцип конструирования “зарядодефицитных”аналогов Spm, заключающийся в замене Spd-фрагмента молекулы на изостерный ему триэтилентетраминовый, поскольку существенное снижение основности вторичных аминогрупп достигается, в том числе, и сокращением расстояния между ними до двух метиленовых звеньев. Аминооксианалоги Spm и Spd (Khomutov A. R., et al. 1996. Tetrahedron, 52, ) принадлежат к другому типу “зарядодефицитных” аналогов полиаминов. Используя эти аналоги, которые, как и все О-замещенные гидроксиламины, являются карбонильными реагентами, были получены оригинальные «бисубстратные» ингибиторы SSAT, хорошо моделирующие промежуточный комплекс Ас-СоА–полиамин. Результаты этих исследований также представлены во второй части данной работы.
Успешное применение химических регуляторов метаболизма во многом определяется возможностью создания форм, активно транспортируемых в клетки. Попытка решения этой проблемы на примере 1-аминоокси-3-аминопропана (АРА), одного из наиболее эффективных ингибиторов орнитиндекарбоксилазы (ODC), действующего на изолированный фермент в nM концентрация, привела к 1-гуанидиноокси-3-аминопропану (GAPA), что стало возможным после того как нами был найден удобный метод гуанидинилирования аминооксигруппы. GAPA активно проникал в амастиготы L.donovani и был эффективен, в отличие от других ингибиторов ODC, и по отношению к солюсурмин-устойчивых формам паразита. Результаты этих исследований представлены в третьей части диссертационной работы.
Синтез рацемических a-метилированных производных спермидина,
N1-ацетилспермидина и спермина.
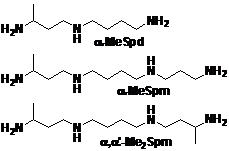 Ингибирование ODC, ключевого фермента биосинтеза полиаминов, приводит к исто-щению пула Spd/Put и торможению роста клеток. Известно, что внесение в культуральную среду как Spd и Spm, так и их a-метилированных производных полностью восстанавливает рост таких клеток (Lakanen J. R., et al. 1992. J.Med.Chem., 35, 724-734). Однако активация катаболизма полиаминов в результате индукции SSAT приводит к истощению внутри-клеточного пула не Put/Spd, а Spm/Spd, поэтому a priori
Ингибирование ODC, ключевого фермента биосинтеза полиаминов, приводит к исто-щению пула Spd/Put и торможению роста клеток. Известно, что внесение в культуральную среду как Spd и Spm, так и их a-метилированных производных полностью восстанавливает рост таких клеток (Lakanen J. R., et al. 1992. J.Med.Chem., 35, 724-734). Однако активация катаболизма полиаминов в результате индукции SSAT приводит к истощению внутри-клеточного пула не Put/Spd, а Spm/Spd, поэтому a priori
Рис. 2. a-Метилированные аналоги полиаминов.
неочевидно, будут ли a-метилполиамины устойчивые к действию SSAT, поддерживать рост клеток с супериндуцированной SSAT, а также устранять в этих условиях патологические последствия дефицита полиаминов in vivo.
Описанные в литературе методы синтеза a-MeSpd, a-MeSpm и a, a’-Me2Spm позволя-ют получать эти соединения в количестве нескольких десятков миллиграммов с низким сум-марным выходом (Lakanen J. R., et al. 1992. J.Med.Chem., 35, 724-734). Проблемы с раствори-мостью ключевых промежуточных соединений, а также трудоемкие процедуры выделения и очистки на последних стадиях синтеза делают нецелесообразным воспроизведение и, тем бо-лее, масштабирование этих методик получения a-метилполиаминов. Соответственно, необ-ходимо было разработать альтернативные схемы синтеза, позволяющие получать a-MeSpd, a-MeSpm и a, a’-Me2Spm с высокими суммарными выходами в количествах, необходимых для проведения экспериментов с лабораторными животными.
Среди разнообразных методов создания C-N-связи, используемых для получения поли-лиаминов (см. обзор: Kuksa V., et al. 2000. Synthesis, 9, ), в настоящей работе для по-строения скелета аналогов полиаминов было использовано алкилирование избытка амина ме-тансульфонатами соответствующих N-защищённых аминоспиртов (Схема 1). Основным ин-термедиатом в синтезе a-MeSpd является N-Cbz-3-аминобутанол (3), который был получен восстановлением LiAlH4 коммерчески доступного этилового эфира 3-аминомасляной кислоты (1) и последующим N-карбобензоксилированием аминоспирта (2). Ключевой стадией синтеза
Схема 1
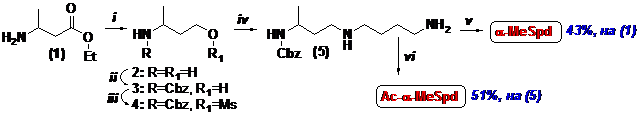
(i)- LiAlH4/THF/Δ; (ii)- CbzCl/THF/H2O/NaHCO3; (iii)- MsCl/Et3N/CH2Cl2; (iv)- H2N(CH2)4NH2/THF;
(v)- (1) H2/Pd/AcOH/MeOH; (2) HCl/MeOH; (vi)- (1) Boc2O; (2) H2/Pd; (3) AcCl/Et3N/THF; (4) HCl/MeOH.
являлось алкилирование 1,4-диаминобутана метансульфонатом (4), которое проводили при 0°С, что позволило уменьшить количество минорных побочных продуктов, трудно отделяе-мых от N-Cbz-a-MeSpd (5) хроматографией на силикагеле. Защитную группу удаляли ката-литическим гидрированием над Pd-чернью и целевой легко кристаллизующийся из водного спирта трихлоргидрат a-MeSpd был получен с суммарным выходом 43%, считая на (1).
Для построения скелета a-MeSpm полиаминную цепь последовательно наращивали алкилируя соответствующие амины мезилатами N-защищенных аминоспиртов (Схема 2), в целом, аналогично описанному выше для a-MeSpd, что позволило получить целевой a-MeSpm в виде хорошо кристаллизующегося тетрахлоргидрата с выходом 41%, считая на (4).
Схема 2
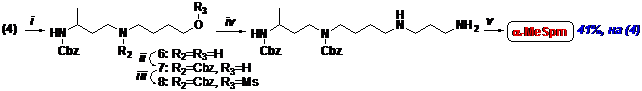
(i)- H2N(CH2)4OH/THF; (ii)- CbzCl/THF/H2O/NaHCO3; (iii)- MsCl/Et3N/CH2Cl2; (iv)- H2N(CH2)3NH2/THF;
(v)- (1) H2/Pd/AcOH/MeOH; (2) HCl/MeOH.
Описанная выше линейная стратегия малоэффективна для получения a, a’-Me2Spm. Симметрия молекулы целевого соединения позволила осуществить конвергентный синтез (Схема 3), ключевой стадией которого являлось алкилирование бис-о-нитрофенилсульфо-нильного (Ns) производного 1,4-диаминобутана (11) N-Cbz-3-аминобутилбромидом (10) и целевой a, w-Me2Spm был получен в виде тетрахлоргидрата с суммарным выходом 57%, считая на (4).
Схема 3
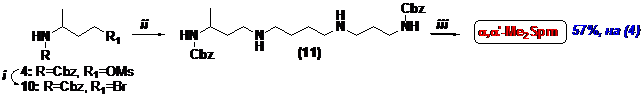
(i)- LiBr/THF; (ii)- (1) NsNH(CH2)4NHNs/DMF/K2CO3; (2) PhSH/DMF/K2CO3; (iii)- (1) H2/Pd/AcOH/MeOH;
(2) HCl/MeOH.
Синтезированные такими способами a-MeSpd, a-MeSpm и a, a’-Me2Spm по данным ВЭЖХ в стандартных условиях анализа полиаминов (Hyvonen T., et al. 1992. J. Chromatogr., 574, 17-21) имели чистоту более 99.5%.
Активность рацемических a-метилированных аналогов Spd и Spm in vivo. [*]
Активация катаболизма полиаминов в результате супериндукции SSAT нетоксичны-ми дозами солей Zn2+ и диэтил-нор-Spm (индуктор SSAT) у SSAT-трансгенных крыс приво-дит к истощению пула Spd и Spm, наиболее выраженному в поджелудочной железе и печени, и развитию острого некротизирующего панкреатита, который вызывает 100% гибель живот-ных в течение 72 ч (Alhonen L., et al. 2000. Proc.Natl.Acad.Sci.USA, 97, ). К моменту начала настоящего исследования не было известно способов обеспечения функциональной целостности поджелудочной железы на фоне активации SSAT.
Оказалось, что даже однократное введение a-MeSpd (50 мг/кг) SSAT-трансгенным крысам до индукции фермента полностью предотвращало развитие острого панкреатита. Более того, введение крысам a-MeSpd или a, a’-Me2Spm через 6 ч после индукции фермента существенно снижает тяжесть заболевания и исключает смертность животных как минимум в течение 14 сут (выживаемость составляет 100% в случае a-MeSpd и 90% в случае a, a’-Me2Spm). Соответственно, именно дефицит полиаминов, по-видимому, является причиной развития панкреатита у SSAT-трансгенных крыс. Следует отметить, что в поджелудочной железе a, a’-Me2Spm эффективно (~ на 35% за 48 ч) расщеплялся до a-MeSpd, что свидетель-ствует о его катаболической неустойчивости in vivo.
Известно, что одним из факторов инициации острого панкреатита является активация трипсиногена, однако молекулярные механизмы этого процесса окончательно не установле-ны (Halangk W., et al. 2002. Am.J.Physiol.Gastrointest.Liver Physiol., 282, 367–374). При обра-ботке солями Zn2+ изолированных ацинарных клеток поджелудочной железы SSAT-транс-генных крыс наблюдается быстрое снижение внутриклеточной концентрации полиаминов в результате индукции SSAT и существенное повышение активности трипсиногена. Предвари-тельное введение животным a, a’-Me2Spm блокирует активацию трипсиногена в ацинарных клетках, что свидетельствует о возможном участии полиаминов в процессах регуляции ак-тивности клеточных протеаз.
Следует отметить, что и на других лабораторных моделях развитие панкреатита со-провождается активацией катаболизма полиаминов в поджелудочной железе, приводящей к истощению пула Spm и Spd. Подобные изменения были отмечены у немногочисленных об-следованных пациентов, страдающих острым или хроническим панкреатитом. Поскольку на-бор веществ, используемых для терапии острого панкреатита, весьма ограничен, можно предположить, что катаболически устойчивые аналоги Spm и Spd представят определенный интерес и для практической медицины.
Регенерация печени крыс после частичной гепатектомии была одним из первых при-меров, на котором in vivo удалось показать необходимость полиаминов для нормального рос-та животных клеток (Janne J. 1967. Acta Physiol.Scand Suppl., 300, 1–71). На ранних стадиях регенерации наблюдается увеличение концентрации Put и некоторое снижение уровня Spd, а количество Spm практически не изменяется. У SSAT-трансгенных крыс частичная гепатэкто-мия сопровождается резким снижением уровня Spd (Alhonen L., et al. 2002. Biochem.J., 362, 149–153), по-видимому, из-за самопроизвольной индукции SSAT в результате операционно-го вмешательства. У таких крыс способность печени к регенерации восстанавливается лишь через 3 сут, когда за счет высокой активности ODC уровень Spd в печени вновь достигает предоперационного значения. Введение SSAT-трансгенным крысам a-MeSpd перед операци-ей снимает блок регенерации печени, что указывает на важность полиаминов для нормально-го протекания регенерационных процессов. Таким образом, казалось, что именно Spd а не Spm необходим для обеспечения процессов регенерации печени крыс. Однако, введение a, a’-Me2Spm SSAT-трансгенным крысам перед частичной гепатэктомией показало, что этот аналог Spm столь же эффективен, как и a-MeSpd. Превращение a, a’-Me2Spm в a-MeSpd в печени, в отличие от поджелудочной железы, происходит лишь в незначительной степени и концентрация a-MeSpd, образовавшегося из a, a’-Me2Spm, оказывается намного ниже необ-ходимой для инициации регенерации. Таким образом, в данном случае Spm и Spd взаимо-заменяемы. Исследование влияния (R,R)- и (S,S)-диастереомеров a, a’-Me2Spm (синтез и био-химические свойства диастереомеров описаны в следующих разделах) на способность пече-ни SSAT-трансгенных крыс к регенерации после частичной гепатэктомии показало, что оба диастереомера одинаково эффективны, т. е. стереоконфигурация аналога несущественна.
Tаким образом впервые был найден способ обращения последствий супериндукции SSAT in vivo, позволяющий предотвращать развитие острого панкреатита у SSAT-трансген-ных крыс и показано, что a-MeSpd и a, a’-Me2Spm представляют собой функционально-ак-тивные миметики Spd и Spm, соответственно. Полученные данные о метаболизме рацемичес-ких форм этих аналогов свидетельствовали о необходимости получения индивидуальных изомеров и диастереоизомеров и детального изучения их взаимодействия с ферментами метаболима полиаминов.
Синтез оптических изомеров a-метилированных полиаминов
Молекулы a-метилполиаминов хиральны, а оптические изомеры известных хираль-ных аналогов полиаминов обладают разной биологической активностью. Так, среди изоме-ров 2,10-дигидрокси-N1,N11-диэтилнорспермина и 2,10-дигидрокси-N1-циклопропилметил-N11-этилнорспермина лишь один диастереомер каждого из веществ оказался супериндукто-ром SSAT (Bergeron R. J., et al. 2001. J. Med. Chem., 44, ). Можно ожидать, что и сте-реоизомеры a-метилполиаминов будут по-разному узнаваться клеточными полиамин-связы-вающими сайтами и проявлять различную биологическую активность.
Учитывая, что структура целевых a-метилполиаминов не слишком сложна, рациональ-нальным подходом представлялось использование коммерчески доступных энантиомерно чистых исходных соединений. Поэтому для построения 3-аминобутанового фрагмента требу-емой конфигурации использовали энантиомеры 2-аминопропанола (12), углеродный скелет которых удлиняли на 1 атом алкилированием цианид-иона мезилатом N-Вос аланинола (14), что приводило к нитрилу (15). Нитрилы (15) восстанавливали LiAlH4 в эфире при -5˚C, что без рацемизации приводило к аминам, которые превращали в о-нитрофенилсульфонильные (Ns) производные (16), служившие универсальными хиральными блоками для синтеза всех оптических изомеров a-метилполиаминов (Схема 4). Для получения энантиомеров a-MeSpd соединения (16) алкилировали N-(4-йодбутил)-фталимидом, что приводило к полностью за-щищенному триамину (17). Удаление Ns-групп приводило к диза-щищенным a-MeSpd, кото-рые очищали хроматографией на силикагеле. После удаления Boc- и фталильной защитных групп целевые (R)- и (S)- изомеры a-MeSpd были выделены в виде трихлоргидратов с сум-марными выходами 43% и 45%, считая на (12а) и (12b), соответственно.
Схема 4
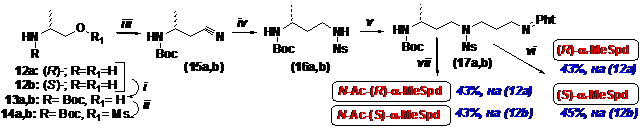
(i)- Boc2O/Et3N/CHCl3; (ii)- MsCl/Et3N/CH2Cl2; (iii)- NaCN/DMSO, 40˚C; (iv)- (1) LiAlH4/Et2O, -5˚C; (2) NsCl/Et3N/CH2Cl2, 0˚C; (v)- PhtN(CH2)4I/K2CO3/DMF; (vi)- (1) PhSH/K2CO3/DMF; (2) N2H4/EtOH/Δ; (3) HCl/MeOH; (vii)- (1) HCl/MeOH; (2) AcCl/Et3N/THF; (3) PhSH/K2CO3/DMF; (4) N2H4/EtOH/Δ; (5) HCl/MeOH.
Для получения (R)- и (S)-a-MeSpm cульфамиды (16) обрабатывали избытком 1-бром-4-хлорбутана в DMF в присутствие поташа (Схема 5). Полученные хлориды превращали в иодиды (18), вводили в реакцию с избытком 1,3-диаминопропана и соединения (19) очи-щали колоночной хроматографией на силикагеле. Следует отметить, что диамины (19) оказа-лись неустойчивы при хранении, вероятно из-за внутри - или межмолекулярного замещения сульфамидной группы. Эти наблюдения вполне согласуются с тем, что Ns-группа легко уда-ляется под действием алкил - и арилсульфид-анионов, нуклеофильность которых значительно выше чем у аминогруппы. Защитные группы в соединениях (19) удаляли последовательной обработкой тиофенолом в DMF и затем раствором HCl в абс. этаноле. Последующая пере-кристаллизация из Н2О-МеОН-EtOH приводила к целевым тетрахлоргидратам (R)- и (S)-a-MeSpm с суммарными выходами 32% и 33%, считая на (12а) и (12b), соответственно.
Схема 5
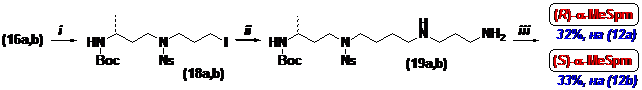
(i)- (1) Cl(CH2)4Br/DMF/K2CO3; (2) NaI/(CH3)2CO; (ii)- H2N(CH2)3NH2/THF; (iii)- (1) PhSH/K2CO3/DMF;
(2) HCl/EtOH.
Молекула a, a’-Me2Spm симметрична и поэтому существуют лишь три диастереомера – (R,R), (S,S) и (R,S). Для получения (R,R)- и (S,S)- изомеров a, a’-Me2Spm сульфамиды (16) вводили в реакцию со стехиометрическим количеством дииодбутана и затем “one-pot” удаля-ли Ns-группы (Схема 6). При этом следы не вошедших в реакцию соединений (16) после де-нозилирования превращались в соответствующие (R)- и (S)-изомеры N3-(трет-бутилоксикар-бонил)-1,3-диаминобутана, которые плохо отделялись хроматографией от ди-Вос-производ-ных (20). Поэтому перед удалением Ns-групп к реакционной смеси прибавляли бензилбромид (10 мольн. %) для алкилирования не вошедших в реакцию с дийодбутаном нозилатов (16). Та-кая последовательность операций позволяла легко выделять в чистом виде ди-Вос-производ-ные (20). После удаления Вос-групп целевые (R,R)- и (S,S)-a, a’-Me2Spm получали в виде тет-рахлоргидратов с суммарными выходами 42% и 46 %, считая на (12а) и (12b), соответственно
Схема 6
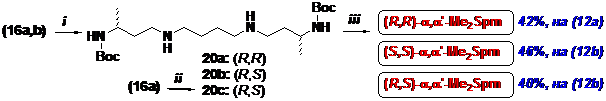
(i)- (1) I(CH2)4I/K2CO3/DMF; (2) PhSH/K2CO3/DMF; (ii)- 17b/K2CO3/DMF; (2) PhSH/K2CO3/DMF; (iii)- HCl/EtOH.
Мезо-форму a, a’-Me2Spm получали из иодида (18b), который обрабатывали стехио-метрическим количеством сульфамида (16a) (Схема 6) аналогично описанному выше для (R,R)- и (S,S)-диастереолмеров и (R,S)-ди-Вос-a, a’-Me2Spm (20c) выделяли хроматографией на силикагеле. Вос-группы удаляли действием HCl в абс. этаноле и целевой (R,S)-a, a’-Me2Spm получали в виде тетрахлоргидрата с суммарным выходом 40 %, считая на (12b).
Синтезированные такими способами неописанные ранее изомеры a-MeSpd и a-MeSpm, а также диастереомеры a, a’-Me2Spm по данным ВЭЖХ в стандартных условиях анализа по-лиаминов (Hyvonen T., et al. 1992. J. Chromatogr., 574, 17-21) имели чистоту более 99.5%.
Взаимодействие оптических изомеров a-метилированных

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
