
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рис. 15. Способность C-метилиро-ванных аналогов Spd (100 µM) поддерживать рост клеток DU145 с истощенным в результате инку-бации с DFMO (5 mM) пулом по-лиаминов. (А) – в присутствии 1 mM AG; (В) – без 1 mM AG.
(Nagarajan S., et al. 1988. Biochem. J., 254, 373-378). Соответственно и С-метилированные аналоги Spd, представляю-щие собой более тонкий инструмент исследования, в 100 μM концентрации в примерно рав-ной степени восстанавливали рост клеток подобно 100 μM Spd (Рис. 15). Таким образом, g-MeSpd представляет собой не только новый метаболически устойчивый, но и функциональ-но-активный миметик Spd, что делает его ценным инструментом исследования, позволяю-щим, в том числе, раздельно изучать клеточные функции Spm и Spd в условиях острого де-фицита полиаминов.
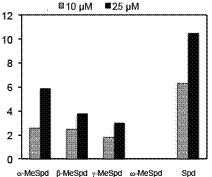
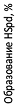 Вместе с тем, в условиях хронического дефицита полиаминов, приводящего к “недоги-пузинилированию” eIF5А (инкубация клеток с DFMO от 12 сут и более), активности аналогов сильно отличались и лишь a - и b-MeSpd были способны восстанавливать рост клеток (Рис.15) Поэтому были исследованы субстратные свойства аналогов в DHS-реакции (Рис. 16). Оказа-лось, что w-MeSpd не является субстратом DHS, тогда как три другиx аналога были донорами аминобутильного остат-ка, хотя их субстратные свойства и ухудшались при переме-щении метильной группы от a- к g-углеродному атому Spd.
Вместе с тем, в условиях хронического дефицита полиаминов, приводящего к “недоги-пузинилированию” eIF5А (инкубация клеток с DFMO от 12 сут и более), активности аналогов сильно отличались и лишь a - и b-MeSpd были способны восстанавливать рост клеток (Рис.15) Поэтому были исследованы субстратные свойства аналогов в DHS-реакции (Рис. 16). Оказа-лось, что w-MeSpd не является субстратом DHS, тогда как три другиx аналога были донорами аминобутильного остат-ка, хотя их субстратные свойства и ухудшались при переме-щении метильной группы от a- к g-углеродному атому Spd.
Рис. 16. Субстратные свойства C-метилированных аналогов Spd в дезоксигипузинсинтазной реакции. Условия реакции см. в подписи к Рис. 5.

 В случае (R)- и (S)-изомеров a-MeSpd (см. пред. раздел) и w-MeSpd (см. выше) наблюдалась прямая корреляция между субстратными свойствами ана-логов в DHS-реакции и способностью преодолевать хро-нический дефицит поли-аминов в клетке, вызванный ин-кубацией с клеток с DFMO от 12 сут и более (Рис. 17). Однако в случае g-MeSpd зависимости оказались более сложными и поэтому возникла необходимость прямо оце-нить количество гипузинилированного eIF5А, возникаю-щего в клетках дефицитных по Spd и обработанных раце-матами C-метилированных ана-логов Spd. Оказалось, что в клетках DU145 из g-MeSpd не образуется гипузинили-рованный eIF5А, что может быть обусловлено тем, что субстратом DHS является непроникающий в клетки изо-мер g-MeSpd.
В случае (R)- и (S)-изомеров a-MeSpd (см. пред. раздел) и w-MeSpd (см. выше) наблюдалась прямая корреляция между субстратными свойствами ана-логов в DHS-реакции и способностью преодолевать хро-нический дефицит поли-аминов в клетке, вызванный ин-кубацией с клеток с DFMO от 12 сут и более (Рис. 17). Однако в случае g-MeSpd зависимости оказались более сложными и поэтому возникла необходимость прямо оце-нить количество гипузинилированного eIF5А, возникаю-щего в клетках дефицитных по Spd и обработанных раце-матами C-метилированных ана-логов Spd. Оказалось, что в клетках DU145 из g-MeSpd не образуется гипузинили-рованный eIF5А, что может быть обусловлено тем, что субстратом DHS является непроникающий в клетки изо-мер g-MeSpd.
Рис. 17. Влияние C-метилированных аналогов Spd на гипузини-лирование eIF5A в клетках с истощенным пулом Spd. Клетки DU145 выращивали в присутствии DFMO (5 mM) и аналогов Spd (100 µM) в течение 9 сут с 1 mM AG, или без AG. Гипузинилирован-ный eIF5A (pI, 5.37) негипузинилированный eIF5A (pI 5.25) и ацети-лированную форму негипузинилированного eIF5A (pI, 5.1) разделяли двумерным электрофорезом и белки визуализировали, используя антитела к eIF5A.
Таким образом, использование системы неизвест-ных ранее C-метилированных аналогов полиаминов открывает неожиданные возможности для прецизионного воздействия на метаболизм полиаминов, а различия в биохимических свойствах аналогов Spm и Spd определяются как положением метильной группы, так и конфигурацией a-углеродного атома. Таким образом, ферменты метаболизма Spm и Spd, AZ-зависимая система регуляции гомеостаза полиаминов и система транспорта полиаминов имеют не только «электростатические», но и дополнительные, различающиеся между собой, структурные критерии узнавания Spm и Spd, используя которые оказывается возможным создавать новые избирательно действующие структуры и исследовать клеточные функции легко взаимопревращающихся и частично взаимозаменяемых Spm и Spd.
Синтез SpmTrien – нового изостерного зарядодефицитного аналога Spm
и его биохимически значимых производных.
Биологические эффекты полиаминов определяются геометрией молекулы, обеспечи-вающей необходимое пространственное расположение аминогрупп и степенью их протони-рования. Большая часть работ по изучению зависимости биологической активности поли-аминов от их строения посвящена изучению структурных аналогов Spm и Spd, включая про-изводные с различными заместителями при концевых атомах азота (см. обзоры: Casero R. A. & Woster P. M. 2009. J. Med. Chem., 52, ; Casero R. A. & Marton L. J. 2007. Nat. Rev. Drug Discov. 6, 373-390). Исследования вклада заряда аминогрупп Spm/Spd в биологические эффекты полиаминов не столь многочисленны. В подобных работах полезным инструментом являются зарядодефицитные аналоги полиаминов, а рациональным подходом к созданию та-ких веществ является понижение основности аминогрупп при минимальном искажении гео-метрии молекулы, что, однако, существенно ограничивает число возможных аналогов.
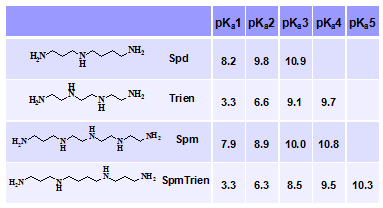 В настоящей работе предложен и реализован оригинальный подход к созданию ново-го зарядодефицитного аналога Spm, состоящий в сокращении расстоя-ния между вторичными аминогруп-пами до 2-х метиленовых звеньев, что существенно понижает основ-ность вторичных аминогрупп ана- лога по сравнению со Spm (Табл. 8).
В настоящей работе предложен и реализован оригинальный подход к созданию ново-го зарядодефицитного аналога Spm, состоящий в сокращении расстоя-ния между вторичными аминогруп-пами до 2-х метиленовых звеньев, что существенно понижает основ-ность вторичных аминогрупп ана- лога по сравнению со Spm (Табл. 8).
Табл. 8. Основность аминогрупп Spd, Trien, Spm и SpmTrien.
Использование 15N-ЯМР спектроскопии для определения микроскопических констант диссоциации показало существование равновесия между трипротонированными формами SpmTrien в интервале рН 6.6-8.5 с протоном, делокализованным между тремя вторичными аминогруппами (Рис. 18). Это свойство может оказаться весьма полезным для обеспечения продуктивного взаимодействия SpmTrien с полиамин-связывающими сайтами, т. к. аналог
способен трансформироваться в ионную форму, комплементарную участку связывания.
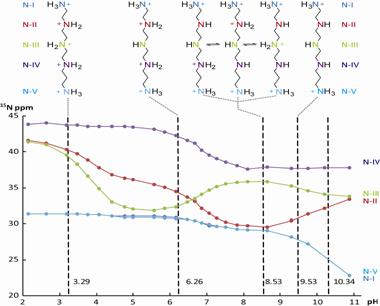 Рис. 18. Зависимость хим. сдвига атомов азота от значения рН в 15N-ЯМР спектрах SpmTrien
Рис. 18. Зависимость хим. сдвига атомов азота от значения рН в 15N-ЯМР спектрах SpmTrien
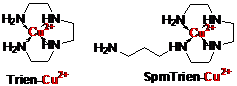 Замена Spd фрагмента молекулы Spm изостерным ему триэтилентетрами-новым приводит к появлению в мо-лекуле SpmTrien сразу трех этилен-диаминовых фрагментов, что дает начало уникальной для биохимии полиаминов способности к комплек-сообразованию с ионами переход-ных металлов (Рис. 19). Известно, что триэтилентетрамин (Trien) обра-зует очень прочный комплекс с ионами Cu2+ [pKuns 20.4 при pH 14 и pKuns 14.2 при pH 7 (Schwarzenbach G. 1950. Helv. Chem. Acta 33, 974-985)], а для комплекса SpmTrien с ионами Cu2+ можно ожидать близких значений констант диссо-циации.
Замена Spd фрагмента молекулы Spm изостерным ему триэтилентетрами-новым приводит к появлению в мо-лекуле SpmTrien сразу трех этилен-диаминовых фрагментов, что дает начало уникальной для биохимии полиаминов способности к комплек-сообразованию с ионами переход-ных металлов (Рис. 19). Известно, что триэтилентетрамин (Trien) обра-зует очень прочный комплекс с ионами Cu2+ [pKuns 20.4 при pH 14 и pKuns 14.2 при pH 7 (Schwarzenbach G. 1950. Helv. Chem. Acta 33, 974-985)], а для комплекса SpmTrien с ионами Cu2+ можно ожидать близких значений констант диссо-циации.
Рис. 19. Cтруктуры комплексов Trien и SpmTrien с ионами Cu2+
Следует отметить, что ни Spm, ни его известные анало-ги не обладают подобным комплексом свойств. Соответственно, SpmTrien может быть ис-пользован в качестве регулятора активности ферментов метаболизма полиаминов, для изуче-ния особенностей активного транспорта Spm в клетки и исследования взаимодействий поли-аминов с нуклеиновыми кислотами. При этом биологические эффекты SpmTrien могут быть как рН-, так и Cu2+-зависимыми.
Выбранная нами стратегия синтеза SpmTrien состояла в последовательном наращива-нии полиаминной цепи путем алкилирования соответствующих аминов мезилатами N-защи-щенных аминоспиртов. Первый способ синтеза предусматривает построение полиаминной цепи, начиная с С-3 фрагмента (Схема 12). Исходным соединением служил 3-аминопропа-
Схема 12
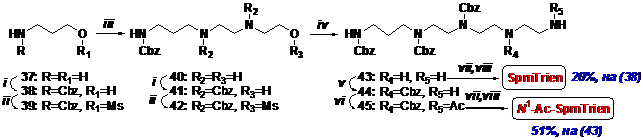
i- CbzCl/NaHCO3/THF/H2O; ii- MsCl/Et3N/CH2Cl2; iii- H2NCH2CH2NHCH2CH2OH/THF; iv- H2NCH2CH2NH2/THF; v- 1. o-C6H4(OH)CHO/THF; 2. i-; 3. MeONH2/THF/H2O; vi- AcCl/Et3N/THF; vii-H2/Pd/AcOH/MeOH; viii- HCl/MeOH
нол-1 (37), который N-карбобензоксилировали, превращали в соответствующий мезилат и затем без выделения вводили в реакцию с избытком N-(2-аминоэтил)-аминоэтанола в THF. Соединение (40) выделяли колоночной хроматографией на силикагеле, но оно содержало в качестве трудно отделяемой примеси небольшое количество продукта алкилирования N-(2-аминоэтил)аминоэтанола по вторичной аминогруппе. Вторичные аминогруппы в соединении (40) защищали и после хроматографии на силикагеле получали чистый трис-Cbz-аминоспирт (41). Его превращали в мезилат, который без выделения обрабатывали избытком этиленди-амина, что приводило к трис-Cbz-диамину (43) из которого получали пентахлоргидрат SpmTrien с выходом 20%, считая на N-Cbz-аминоспирт (38) и тетрахлоргидрат N1-AcSpmTrien с выходом 51%, считая на соединение (43).
Второй путь синтеза SpmTrien предусматривает построение полиаминной цепи, начи-ная с С-2 фрагмента (Схема 13), что позволяет избежать неоднозначно протекающей стадии алкилирования первичной аминогруппы в присутствии незащищенной вторичной.
Схема 13
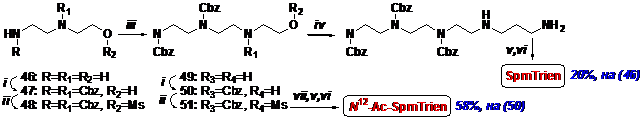
i- CbzCl/NaHCO3/THF/H2O; ii- MsCl/Et3N/CH2Cl2; iii- H2NCH2CH2OH/THF; iv- H2NCH2CH2CH2NH2/THF; v- H2/Pd/AcOH/MeOH; vi- HCl/MeOH; vii- H2N(CH2)3NHAc/THF.
Методически этот путь аналогичен первому и представляет собой семистадийный процесс. Однако выделение и очистка промежуточных соединений менее трудоемки. Суммарный вы-ход SpmTrien составил 20%, считая на N-(аминоэтил)аминоэтанол, а N12-AcSpmTrien был по-лучен в виде тетрахлоргидрата с выходом 58%, считая на трис-Cbz-аминоспирт (50).
Таким образом были синтезированы SpmTrien и два неизвестных ранее моно-ацетиль-ных производных, представляющих интерес для исследования SSAT и АPAO.
Терминально бис-алкилированные аналоги полиаминов являются одним из основных инструментов исследования системы метаболизма полиаминов и, кроме того, имеют опреде-ленные перспективы клинического применения. Тем не менее, механизмы их действия на молекулярном уровне в большинстве случаев до сих пор неизвестны. Влияние геометрии молекулы на биологические эффекты бис-алкилполиаминов подробно исследовалось на протяжении последних 15 лет (см. обзоры: Casero R. A. & Woster P. M. 2009. J. Med. Chem., 52, ; Casero R. A. & Marton L. J. 2007. Nat. Rev. Drug Discov. 6, 373-390; Casero R. A. & Woster P. M. 2001. J.Med.Chem., 44, 1-26), но вклад зарядовой составляющей в биологическую активность практически не изучен.
Для получения неизвестного ранее зарядодефицитного N1,N12-Et2SpmTrien нами был
использован N-этиламиноэтанол, который карбобензоксилировали, спирт (53) превращали в мезилат и без очистки вводили в реакцию с избытком N-(2-аминоэтил)аминоэтанола (Схема 14). Диаминоспирт (55), содержащий небольшое количество изомерного продукта алкилиро-вания N-(2-аминоэтил)аминоэтанола по вторичной аминогруппе, вновь карбобензоксилиро-вали. При этом целевое трис-Cbz производное (56) легко отделялось хроматографией на си-ликагеле от побочного продукта реакции алкилирования, содержащего две Cbz-группы. Защи-щённый аминоспирт (56) превращали в мезилат (57), который обрабатывали избытком N-этил-1,3-диаминопропана. Наряду с целевым соединением линейного строения (58) образовывалось некоторое количество продукта алкилирования N-этил-1,3-диаминопропана по вторичной ами-ногруппе, который плохо отделялся хроматографией. Для удаления примеси сырой диамин (58) обрабатывали ~20 мольн. % салицилового альдегида, что приводило к образованию устой-чивого основания Шиффа по первичной аминогруппе примеси, которое легко отделялось хро-матографией от основного продукта (58). После удаления Cbz-защитных групп N1,N12-Et2-SpmTrien был получен в виде пентахлоргидрата с суммарным выходом 8%, считая на (52).
Схема 14

i- CbzCl/THF/H2O/NaHCO3; ii- MsCl/Et3N/CH2Cl2; iii- H2NCH2CH2NHCH2CH2OH/THF; iv- C2H5NH(CH2)3NH2/THF; v- H2/Pd/MeOH/AcOH; vi- HCl/MeOH.
Таким образом, на основе скелета SpmTrien был получен первый зарядодефицитный по центральному фрагменту аналог бис-этилспермина – N1,N12-Et2-SpmTrien.
Взаимодействие SpmTrien и его производных с ферментами метаболизма
полиаминов и клетками.
Триэтилентетрамин (Trien), является эффективным хелатором ионов Cu2+ (Рис. 19) и используется в практической медицине для лечения болезни Вильсона (Dixon H. B.F., et al. 1972. Lancet, pp. 853–854), обусловленной нарушением обмена меди в организме. В послед-ние годы показана перспективность Trien для снижения гипертрофии левого желудочка, характерной для поздних стадий диабета II типа (Cooper G. J., et al. 2004. Diabetes, 53, ; Cooper G. J., et al. 2009. Diabetologia, 52, 715–722). В настоящее время Trien находится на II стадии клинических испытаний и его метаболизм детально исследуется (Lu J., et al. 2010 J.Clin.Pharmacol., 50, 647–658). Однако, участие системы транспорта полиаминов в доставке Trien в клетки, равно как и участие ферментов метаболизма полиаминов во внутриклеточных трансформациях Trien не рассматривается, несмотря на то, что Trien является изостерным, хотя и зарядодефицитным, аналогом Spd (Табл. 8). Поэтому в этой части работы мы исследо-вали взаимодействие Trien и SpmTrien с ферментами метаболизма полиаминов и клеточные эффекты этих аналогов.
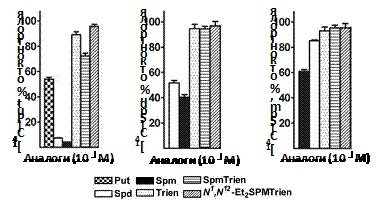 Оказалось, что Trien и SpmTrien, несмотря на структурное соответствие Spd и Spm, не конкурируют с полиаминами за про-никновение в клетки DU145 и, скорее все-го, попадают в клетки не исполь-зуя систему транспорта полиаминов (Рис. 20).
Оказалось, что Trien и SpmTrien, несмотря на структурное соответствие Spd и Spm, не конкурируют с полиаминами за про-никновение в клетки DU145 и, скорее все-го, попадают в клетки не исполь-зуя систему транспорта полиаминов (Рис. 20).
Рис. 20. Конкуренция Trien, SpmTrien и N1,N12-Et2-SpmTrien с полиаминами в процессе транспорта в клетки DU145.
Таким образом, “правильное прото-нирование” аналога оказывается критическим для его узнавания системой активного транс-порта полиаминов.
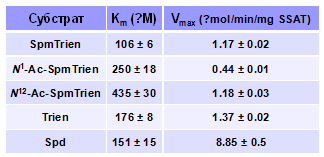 Известными метаболитами Trien являются N1-Ac-Trien и N1,N8-Ac2-Trien, которые вместе с Trien были обнаружены в моче людей принимавших это лекарство (Lu J., et al. 2007. Drug Metab.Dispos., 35, 221–227). Учитывая структурное сходство Trien и SpmTrien с Spd и Spm, соответственно, мы исследовали взаимодействие этих аналогов с SSAT (Табл. 10). Ока-залось, что Trien и SpmTrien являются субстратами этого фермента, но худшими чем Spd, а каждый из продуктов моно-ацетилирования SpmTrien, подобно N1-Ac-Trien, в свою очередь оказался плохим субстратом SSAT. Таким образом, SSAT может рассматриваться в ка-честве одного из ферментов ответственных за ацетилирование Trien in vivo.
Известными метаболитами Trien являются N1-Ac-Trien и N1,N8-Ac2-Trien, которые вместе с Trien были обнаружены в моче людей принимавших это лекарство (Lu J., et al. 2007. Drug Metab.Dispos., 35, 221–227). Учитывая структурное сходство Trien и SpmTrien с Spd и Spm, соответственно, мы исследовали взаимодействие этих аналогов с SSAT (Табл. 10). Ока-залось, что Trien и SpmTrien являются субстратами этого фермента, но худшими чем Spd, а каждый из продуктов моно-ацетилирования SpmTrien, подобно N1-Ac-Trien, в свою очередь оказался плохим субстратом SSAT. Таким образом, SSAT может рассматриваться в ка-честве одного из ферментов ответственных за ацетилирование Trien in vivo.
Табл.10. Взаимодействие Trien, SpmTrien и его моно-ацетильных производных с SSAT.
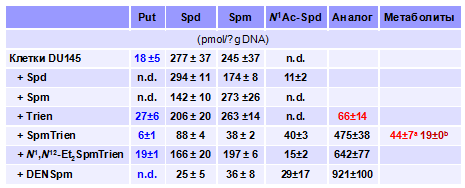 Инкубация клеток DU145 с SpmTrien и Trien и анализ пула полиаминов показали, что моно-ацетильное производное (N1-Ac-SpmTrien) воз-никает лишь из первого аналога, а Trien в клетках DU145 заметно не метаболизирует (Табл. 11) Возможно это связано с тем, что уровень SpmTrien и его метаболитов в клетках почти в 10 раз выше чем содержание Trien и количество N1-Ac-Trien оказывается ниже детектируемо-го уровня. Следует особо отметить, что SpmTrien метаболизирует в клетке и до Trien, причем коли-
Инкубация клеток DU145 с SpmTrien и Trien и анализ пула полиаминов показали, что моно-ацетильное производное (N1-Ac-SpmTrien) воз-никает лишь из первого аналога, а Trien в клетках DU145 заметно не метаболизирует (Табл. 11) Возможно это связано с тем, что уровень SpmTrien и его метаболитов в клетках почти в 10 раз выше чем содержание Trien и количество N1-Ac-Trien оказывается ниже детектируемо-го уровня. Следует особо отметить, что SpmTrien метаболизирует в клетке и до Trien, причем коли-
Табл.11. Метаболизм Trien и SpmTrien в клетках DU145. Влияние аналогов на содер-жание полиаминов в клетках
![]()
чество последнего, возникающего из SpmTrien (концентрация в среде 50 mM), сравнимо с ко-личеством Trien, проникающего в клетку в результате инкубации с 50 mM аналога (Табл. 11). Образование Trien в клетке происходит, по-видимому, через N12-Ac-SpmTrien, который под действием АРАО превращается в Trien. Поэтому из двух моно-ацетильных производных SpmTrien (SSAT, оказывается, не способна дискриминировать аминопропильный и амино-этильный концы молекулы SpmTrien) в клетке детектируется лишь N1-Ac-SpmTrien, не явля-ющийся субстратом АРАО. Таким образом, SpmTrien, накапливается в клетках почти в 10 раз эффективнее Trien, что, учитывая высокую прочность комплексов SpmTrien–Cu2+, может представлять интерес и для практической терапии.
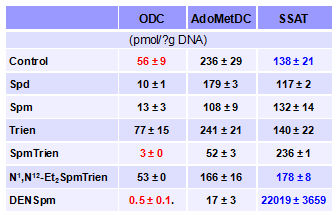 SpmTrien, Trien и N1,N12-Et2-SpmTrien по-разному влияют на внутриклеточное содер-жание полиаминов в клетках DU145 (Табл. 11). Если Trien и N1,N12-Et2-SpmTrien, в особен-ности первый, малоактивны, то SpmTrien эффективно снижал внутриклеточный уровень по-лиаминов, активируя ферменты их катаболизма (образование значительных количеств N1-AcSpd) и подавляя систему их биосинтеза (сильное снижение уровня Put). Наблюдаемые из-менения в активностях ODC, AdoMetDC и SSAT в клетке подтверждали эти предположение (Табл. 12). Следует отметить, что зарядодефицитный N1,N12-Et2-SpmTrien, в отличие от изо-стерного ему DENSpm, не был индуктором SSAT, по-видимому из-за невозможности пра-вильного связывания с С-концевой последовательностью фермента, что и стабилизирует бе-лок, снижая скорость его протеолиза. При этом, N1,N12-Et2-SpmTrien и DENSpm при-мерно одинаково индуцируют продуктив-ный сплайсинг пре-мРНК SSAT (данные не приведены).
SpmTrien, Trien и N1,N12-Et2-SpmTrien по-разному влияют на внутриклеточное содер-жание полиаминов в клетках DU145 (Табл. 11). Если Trien и N1,N12-Et2-SpmTrien, в особен-ности первый, малоактивны, то SpmTrien эффективно снижал внутриклеточный уровень по-лиаминов, активируя ферменты их катаболизма (образование значительных количеств N1-AcSpd) и подавляя систему их биосинтеза (сильное снижение уровня Put). Наблюдаемые из-менения в активностях ODC, AdoMetDC и SSAT в клетке подтверждали эти предположение (Табл. 12). Следует отметить, что зарядодефицитный N1,N12-Et2-SpmTrien, в отличие от изо-стерного ему DENSpm, не был индуктором SSAT, по-видимому из-за невозможности пра-вильного связывания с С-концевой последовательностью фермента, что и стабилизирует бе-лок, снижая скорость его протеолиза. При этом, N1,N12-Et2-SpmTrien и DENSpm при-мерно одинаково индуцируют продуктив-ный сплайсинг пре-мРНК SSAT (данные не приведены).
Табл.12. Влияние полиаминов, DENSpm и заря-додефицитных аналогов Spm на активность фер-ментов метаболизма Spm и Spd клетках DU145. Концентрация полиаминов и их аналогов - 50 mM. Клетки выращивали в присутствии 1 mM AG.
Вызываемое SpmTrien снижение активности ODC оказалось AZ-опосредованным. Из всех аналогов транспорт SpmTrien в большей степени контролировался уровнем AZ – обра-ботка клеток СНХ (ингибитор биосинтеза белка) приводила к резкому увеличению содержа-ния SpmTrien в клетке, тогда как эффекты СНХ на транспорт N1,N12-Et2-SpmTrien и Trien от-сутствовали вовсе, а в случае DENSpm были менее выраженными (Рис. 21). Оказалось, что SpmTrien способен подобно природным полиаминам и многим их аналогам индуцировать +1 сдвиг рамки считывания мРНК AZ, что необходимо для биосинтеза активного белка. Соот-ветственно, количество синтезирующегося в клетке AZ, в ответ на инкубацию с аналогами сильно отличалось, а среди зарядодефицитных производных SpmTrien оказался наиболее ак-тивным (Рис. 21).
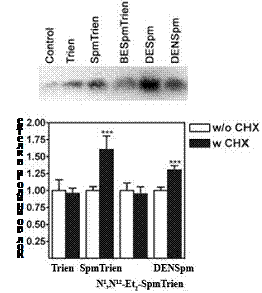 Рис. 21. Влияние Trien, SpmTrien и N1,N12-Et2-SpmTrien на биосинтез AZ (a). Зависимость транспорта аналогов от уровня AZ в клетках DU145 (b). (a) Клетки инкубировали 4 ч с исследуемыми аналогами (50 mM); аликвоту суммарной белковой фракции (50 mg) анализировали электрофорезом в 12% SDS-полиакриламидном геле и визуализировали при помощи антител к AZ. (b) Клетки инкубировали 1 ч с CHX (10 mg/mL), или без CHX, а затем еще 4 ч с Trien, SpmTrien, N1,N12-Et2-SpmTrien и DENSpm (50 mM).
Рис. 21. Влияние Trien, SpmTrien и N1,N12-Et2-SpmTrien на биосинтез AZ (a). Зависимость транспорта аналогов от уровня AZ в клетках DU145 (b). (a) Клетки инкубировали 4 ч с исследуемыми аналогами (50 mM); аликвоту суммарной белковой фракции (50 mg) анализировали электрофорезом в 12% SDS-полиакриламидном геле и визуализировали при помощи антител к AZ. (b) Клетки инкубировали 1 ч с CHX (10 mg/mL), или без CHX, а затем еще 4 ч с Trien, SpmTrien, N1,N12-Et2-SpmTrien и DENSpm (50 mM).
Таким образом, сокращение расстояния между ами-ногруппами до двух метиленовых звеньев в случае Trien (изостер Spd) приводит к тому, что аналог, все еще являясь субстратом SSAT, перестает узнаваться клеткой как Spd: (i) Trien не использует систему транспорта полиаминов для проникновения в клетку, (ii) не влияет на пул полиаминов в клетке и на активности соответствующих ферментов мета-болизма, (iii) не индуцирует ни биосинтез AZ, (iv) ни продуктивный сплайсинг пре-мРНК SSAT. Превращение Trien в SpmTrien приводит к зарядодефицитному изостеру Spm, кото-рый по большинству биохимических параметров, за исключением механизма проникновения в клетку, может считаться полиамином. Учитывая, что внутриклеточное содержание SpmTrien почти в 10 раз выше чем содержание Trien, а также возможность биосинтеза Trien из SpmTrien и низкую токсичность аналога для клеток, можно предположить, что SpmTrien представляет собой не только интересный инструмент исследования метаболизма и клеточ-ных функций полиаминов, но и не лишен некоторой практической значимости.
Конденсация ДНК под действием SpmTrien.[§]
Взаимодействие с ДНК является одной из основных клеточных функций Spm и Spd. Конденсация ДНК, сопровождающаяся появлением оптической плотности при λ > 320 нм из-за рассеяния падающего УФ-излучения на формирующихся компактных частицах, пред-ставляет собой одну из простейших моделей для оценки эффективности взаимодействия по-лиаминов с ДНК. Способность полиаминов конденсировать ДНК зависит от количества по-ложительных зарядов в молекуле и уменьшается в ряду пентамин > тетрамин > триамин (Saminathan M., et al., 1999. Biochemistry, 38, ).
Основность внутренних аминогрупп SpmTrien понижена, что приводит к недостаточному, по сравнению со Spm, протонированию центрального фрагмента молекулы при физиологичес-ком рН (см. Табл. 8). В то же время в области слабокислых рН степень протонирования SpmTrien соответствует ионному состоянию Spm. Поэтому мы провели исследование ДНК-конденсирующих свойств Spd, Spm и SpmTrien при нейтральных и слабокислых рН, исполь-зуя в качестве критерия концентрацию полиамина (скр), начиная с которой А340 быстро рас-тет с увеличением концентрации полиамина (Рис. 22). При рН 6.8 скр Spm и SpmTrien отлича-ются более, чем в 2,5 раза (25 и 67 mM, соответственно), а скр Spd, который подобно SpmTrien является трикатио-ном при данном рН, составляет уже 420 mM (Рис. 22). Однако при рН 5.5 скр Spm и SpmTrien практически совпадают (22 и 24 mM, соответственно), а скр Spd оказыва-ется равной 300 mM.
 Рис. 22. Зависимость конденса-ции ДНК (A340) от концентрации Spd, Spm и SpmTrien при разных pH. (1) - Spm, pH 6.8; (2) - SpmTrien, pH 6.8; (3) - Spd, pH 6.8; (4) - Spm, pH 5.5; (5) - SpmTrien, pH 5.5; (6) - Spd, pH 5.5.
Рис. 22. Зависимость конденса-ции ДНК (A340) от концентрации Spd, Spm и SpmTrien при разных pH. (1) - Spm, pH 6.8; (2) - SpmTrien, pH 6.8; (3) - Spd, pH 6.8; (4) - Spm, pH 5.5; (5) - SpmTrien, pH 5.5; (6) - Spd, pH 5.5.
Таким образом, при рН 5.5 в процесс конденсации включа-ется четвертая протонирован-ная аминогруппа SpmTrien, что подтверждает правомерность рассмотрения его в качестве за-рядо-дефицитного аналога Spm при нейтральном рН. Следует особо отметить, что при рН 6.8 конденсирующая способность трижды протонированного SpmTrien намного превосходит та-ковую для трикатиона Spd. Наблюдаемые отличия обусловлены или геометрией молекулы (расстоянием между положительными зарядами), или же участием непротонированных вто-ричных аминогрупп SpmTrien во взаимодействии с ДНК, возможно, и в результате образова-ния системы водородных связей.
Известно, что ДНК, сконденсированная под действием Spm или SpmTrien, вновь рас-творяется при добавлении гепарина – полианиона, который эффективно извлекает полиами-ны из комплекса с ДНК (, неопубликованные данные). Поскольку SpmTrien является хорошим хелатором Cu2+, то введение в систему солей меди должно вызывать гепа-риноподобные эффекты благодаря образованию прочного комплекса Cu2+-SpmTrien.
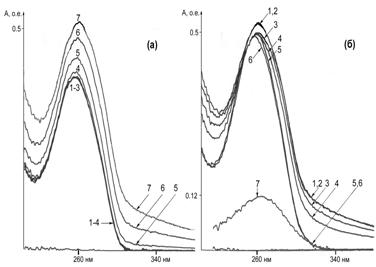 Сравнение УФ-спектров исходной ДНК и ДНК, сконденсированной под действием 19.4 mМ SpmTrien при рН 5.2 (Рис. 23а, кривые 1 и 7) с УФ-спектром конденсированной ДНК после прибавления 19.5 mМ CuCl2 показывает, что в присутствии ионов
Сравнение УФ-спектров исходной ДНК и ДНК, сконденсированной под действием 19.4 mМ SpmTrien при рН 5.2 (Рис. 23а, кривые 1 и 7) с УФ-спектром конденсированной ДНК после прибавления 19.5 mМ CuCl2 показывает, что в присутствии ионов
Рис. 23. Конденсация ДНК под действием SpmTrien (а) и деконденсация в присутст-вии ионов Cu2+ (б). (а): (1) ДНК - 20 mg/ml в 5 mМ Na-Ac буфере, рН 5.2; (2) - (1) + 3.24 mМ SpmTrien; (3) - (1) + 6.48·mМ SpmTrien; (4) - (1) + 9.71·mМ SpmTrien; (5) - (1) + 12.9 mМ SpmTrien; (6) - (1) + 16.2·mМ SpmTrien; (7) - (1) + 19.4·mМ SpmTrien. (б): (1) ДНК - 20 mg/ml в 5 mМ Na-Ac буфере, рН 5.2 + 19.4 mМ SpmTrien; (2) - (1) + 4.96·mМ CuCl2; (3) - (1) + 9.88·mМ CuCl2; (4) - (1) + 14.7·mМ CuCl2; (5) - (1) + 19.5·mМ CuCl2; (6) - (1) + 24.3·mМ CuCl2; (7) 19.4·mМ SpmTrien + 19.5· mМ CuCl2.
Cu2+ происходит растворение компактной формы ДНК (Рис. 23б, кривые 2-6), тогда как в случае ДНК сконденсированной под действием Spm, ионы Cu2+ не вызывают подобного эф-фекта (данные не приведены). В отличие от гепарин-зависимой деконденсации ДНК (УФ-спектры исходной ДНК и комплексов ДНК-Spm и ДНК-SpmTrien после обработки гепари-ном идентичны), в случае ионов Cu2+ величина А260 уменьшается лишь незначительно, что обусловлено заметным поглощением комплекса (Cu*SpmTrien)2+ при 260 нм (Рис. 23б, кри-вая 7). Сами по себе ионы Cu2+ в концентрациях до 2.0 mМ не вызывают изменений УФ-спектра растворов ДНК. Таким образом, наличие у SpmTrien хороших комплексообразую-щих свойств позволяет регулировать его конденсирующую активность не только при помо-щи рН, но и вводя в систему ионы Cu2+, что неизвестно ни для одного из аналогов Spm.
Новые ингибиторы спермин/спермидин N1-ацетилтрансферазы на основе стабильных аналогов бисубстратного комплекса,
возникающего в ходе ферментативной реакции.
Одной из традиционных задач химической энзимологии полиаминов является разра-ботка методов истощения внутриклеточного пула Spm и Spd, что вполне естественно, учиты-вая повышенное содержание полиаминов в опухолевых клетках. Соответственно, были най-дены эффективные индукторы SSAT, ключевого фермента катаболизма полиаминов, однако набор ингибиторов SSAT, в отличие от ODC и AdoMetDC, весьма ограничен. Развитие моле-кулярной биологии полиаминов показало, что полиамин-зависимый сплайсинг пре-мРНК SSAT играет важную роль в регуляции активности фермента, а следовательно и уровня Spm и Spd в клетке (Hyvonen M. T., et al. 2006. RNA, 12, ). Поскольку избирательные ин-гибиторы SSAT являются удобным инструментом исследования активных и неактивных форм фермента, в том числе и методами рентгеноструктурного анализа, представлялось целесообразным расширить существующий набор ингибиторов.
Механизм SSAT-реакции не предусматривает промежуточного образования ацетили-рованного фермента – происходит прямой перенос ацетильной группы Ас-СоА на полиамин. Поэтому для ингибирования фермента были использованы конъюгаты HS-СоА со Spm или Spd, в которых HS-СоА и полиамин были соединены при помощи ацетатного линкера (Рис. 24). Синтезированные производные симметричных полиаминов ингибировали изолирован-ный фермент с IC50 0.5-5.0 mМ в зависимости от строения полиаминного фрагмента (Ervin B. G., et al. 1984. Biochemistry, 23, ), а для получения конъюгатов SH-CoA, а также
|
|
|
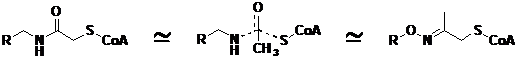
Рис.24. Моделирование переходного состояния бисубстратного комплекса, возникающего в ходе SSAT-реакции, с помощью конъюгатов CoA-полиамин с ацетатным и ацетоновым линкерами.
D-пантотеина, избирательно по N1- или N8-положениям Spd позднее был разработан доста-точно сложный многостадийный синтез (Roblot G. et al. 1993. Tetrahedron, 29, ).
Располагая набором аминооксианалогов Spm и Spd (Khomutov A. R. et al. 1996. Tetra-hedron, 52, ), которые представляют собой не только зарядодефицитные изостер-ные аналоги полиаминов (рКа H2NO-группы ~ 3.5), но являются еще и классическими карбо-нильными реагентами, естественным представлялось использовать различия в реакционной способности H2NO - и H2N-групп для получения конъюгатов CoA-полиамин. На первой ста-дии разработанного нами простого двухстадийного синтеза HS-CoA или D-пантотеин алки-лировали хлорацетоном в слабощелочной среде (Схема 15). Полученный кетон без выделе-ния вводили в реакцию с соответствующим аминооксианалогом Spm или Spd, что приводило к целевым соединениям с выходами, близкими к количественным.
Схема 15.
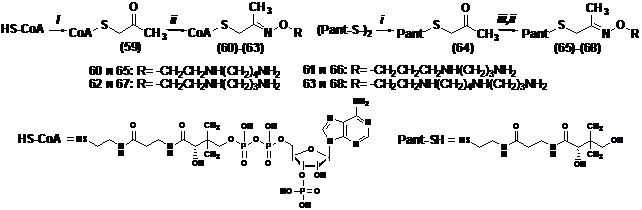
i - ClCH2COCH3/CH3CN/H2O/NaHCO3/Na2CO3; ii - RONH2; iii - DTT/EtOH/H2O
Как следует из данных Табл. 13, наиболее активным оказалось соединение (61), моде-лирующее аддукт по N8-положению Spd, тогда как соединение (60), моделирующее N1-заме-щенный Spd, было не столь эффективно (значения IC50 получены при 4.5 mМ концентрации Ас-СоА в субстратной смеси). Соединение (61) было в 22 раза активнее кетона (59), действо-вавшего хуже любого из CoA-содержащих конъюгатов. Подобная зависимость действия от строения подтверждает вклад полиаминного фрагмента в эффективность торможения. Вклад аденозинового фрагмента СоА в эффективность торможения SSAT был определяющим, что прямо следует из низкой активности пантотеиновых производных имевших IC50 > 100 μМ (данные не приведены). Однако известно, что в случае сукцинил-СоА-ацетоацетат трансферазы пантотеновая часть субстрата вносит значитель-ный вклад в эффективность связывания субстра-та (Fierke C. A. & Jencks W. P. 1986. J.Biol.Chem., 261, )].
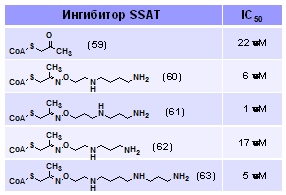 Табл. 13. Ингибирование (IC50) SSAT конъюгатами CoA-SH и D-пантотеина с аминооксианалогами поли-аминов.
Табл. 13. Ингибирование (IC50) SSAT конъюгатами CoA-SH и D-пантотеина с аминооксианалогами поли-аминов.
Таким образом, был получен набор новых аналогов бисубстратного комплекса SSAT-реакции на основе CoA-SH и D-пантотеина и показано, что активность соединений (61), (63) и (60) была сравнима с таковой для лучших из известных ингибиторов SSAT.
Синтез и биологическая активность
функционально-замещенных алкоксигуанидинов.
Среди природных производных гидроксиламина наиболее известны аминокислота ка-наванин (α-амино-γ-гуанидинооксимасляная кислота), широко распространенная в растени-ях, и антибиотик циклосерин (4-аминоизоксазолидон-3), применяемый для лечения туберку-леза. Если синтез и химия циклосерина были разработаны еще в конце 50-х – начале 60-х гг., то удобных способов получения алкоксигуанидинов, в том числе и канаванина, практически не существует. Известно лишь, что обработка защищенного производного каналина S-метил-изотиомочевиной при 20°С в течение 72 ч приводит к канаванину с выходом 10% (Rosenthal G. A. et al. 1983. Anal.Biochem., 133, 277-282), а 5 ч кипячение с цианамидом в спирте позво-ляет получить канаванин с выходом 70% (Frankel M., et al. 1963. J.Chem.Soc., ). Предполагая использовать функционально-замещенные алкоксигуанидины в качестве инструмента исследования метаболизма полиаминов, мы были вынуждены сначала найти удобный метод синтеза этих малодоступных соединений.
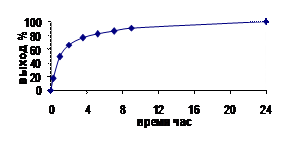
В настоящей работе для получения гуанидинооксиалканов впервые был использован трифлат ди-Boc-гуанидина, который быстро и практически количественно превращает али-фатические амины в соответствующие гуанидины (Feichtinger K., et al. 1998. J.Org.Chem., 1998, 63, ). Однако О-замещенные гидроксиламины существенно менее нуклео-фильны по сравнению с соответствующими алифатическими аминами и поэтому мы сначала исследовали взаимодействие трифлата ди-Boc-гуанидина с модельным О-бензилгидроксил-амином. Оказалось, что ди-Вос-производное бензилоксигуанидина образуется с практически количественным выходом, но реакция протекает существенно медленнее (Рис. 25), чем в слу-чае бензиламина. Величина константы скорости реакции второго порядка (мольные соотно-шения О-бензилгидроксиламина и трифлата ди-Вос-гуанидина), определенная с использова-
зованием 1Н-ЯМР составила 2.66·10-2 л/моль/мин (t1/2 75 мин при 37°С).
![]()
Рис.25. Кинетика гуанидинилирования О-бензилгидрок-силамина (0.3 М) трифлатом ди-Boc-гуанидина (0.3 М) в присутствии Et3N (0.3 М) в CDCl3 при 37°С.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
