
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На следующем этапе работы в реакцию с трифла-том ди-Boc-гуанидина были введены некоторые функционально-замещенные эфиры гидрок-силамина (Схема 16), гуанидилирование которых удается осуществить с высоким выходом несмотря на наличие в радикале меркапто - и гидроксильных групп, а также экзоциклической аминогруппы аденозина.
Схема 16
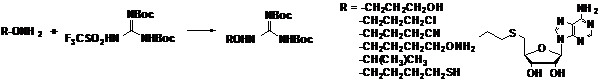
Таким образом, был найден эффективный способ гуанидилирования аминооксигруппы, позволяющий получать целевые гидроксиламин-содержащие аналоги Agm.
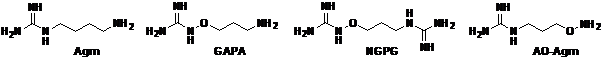
Рис.26. Агматин и его гидроксиламин-содержащие аналоги.
Исходным соединением в синтезах AO-Agm, GAPA и NGPG (Рис. 26) служил 1-эток-сиэтилиденаминоокси-3-аминопропан (69) (Схема 17). Гуанидинилирование его свободной аминогруппы трифлатом ди-Boc-гуанидина позволило получить ди-Boc-производное (69) с выходом близким к количественному. Последующее удаление защитных групп действием HCl/EtOH привело к медленно смокающему на воздухе дихлоргидрату AO-Agm, который был переведен в хорошо кристаллизующийся дитозилат (выход 75.5%, считая на (69)).
Схема 17

i- (BocNH)2C=NTf/Et3N/CH2Cl2/20°C; ii- HCl/i-PrOH/H2O; iii- CbzCl/Et3N/THF; iv- HCl/EtOH/H2O; v- (BocNH)2C=NTf/Et3N/CHCl3/37°C; vi- HBr/AcOH.
Следует отметить, что AO-Agm является изостерным зарядодефицитным аналогом Agm и образует, подобно другим О-замещенным гидроксиламинам, стабильные оксимы с альдегидами и кетонами, включая PLP, который служит коферментом ArgDC. Так как ами-нооксианалоги субстратов пируват - и PLP-зависимых ферментов метаболизма аминокислот являются их эффективными ингибиторами, то следует ожидать высокой активности AO-Agm по отношению к ArgDC.
При получении GAPA свободную аминогруппу соединения (69) сначала карбобензок-силировали, а затем удаляли этоксиэтилиденовую защиту, что приводило к соответствующе-му гидрохлориду (71) (Схема 17). Гуанидинилирование аминооксигруппы трифлатом ди-Boc-гуанидина позволило получить с высоким выходом соединение (72), что после одновремен-ного удаления Boc - и Cbz-защитных групп привело к кристаллическому дибромгидрату GAPA (выход 48.7%, на (69)). Мы реализовали несколько схем получения NGPG, наиболее удачной из которых является синтез, исходящий из 1-аминоокси-3-аминопропана, свободные амино - и аминоокси-группы которого гуанидилировали 1.9 экв. трифлата ди Boc-гуанидина при 37°С (Схема 17). Соединение (73) очищали колоночной хроматографией на силикагеле, и после удаления Boc-защитных групп получали дибромгидрат NGPG (выход 54%, на (69)).
Основность гуанидиноокси-фрагмента GAPA и NGPG имеет значение для правильно-го использования этих аналогов в биохимии полиаминов. Значения pKa аминооксигруппы
![]()
![]()
![]()
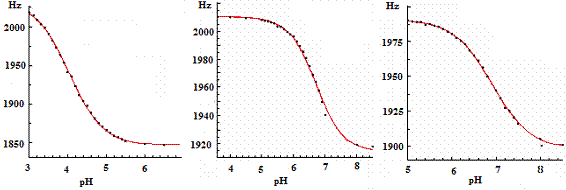
Рис.27. Основность гидроксиламин-содержащих аналогов Agm.
AO-Agm (pKa 4.05) и гуанидинооксигрупп GAPA (pKa 6.7) и NGPG (pKa 6.9) были опреде-лены с использованием 1H-ЯМР спектроскопии (Рис. 27). Таким образом, GAPA, являясь изостером Agm (pKa 9.6 и 12.5), по основности ближе к Put и, скорее всего, будет активно переноситься в клетки, используя, подобно Agm, систему транспорта Put.
Влияние GAPA на размножение Leishmania donovani[**].
Полиамины играют важную роль и в жизненном цикле протозойных паразитов, на-
пример Plasmodium falciparum, Leishmania donovani, Trypanosoma brucei gambiense, вызыва-ющих весьма распространенные в тропиках и субтропиках заболевания – малярию, лейшма-ниоз и сонную болезнь, соответственно (см. обзор: Heby O., et al. 2007. Amino Acids, 33, 359-366). В последние годы возникли формы L.donovani, устойчивые к противолейшманиозным препаратам, в том числе и к широко используемому Cолюсурьмину®, что делает весьма ак-туальным поиск метаболических мишеней, перспективных для создания новых средств борь-бы с лейшманиозом (см. обзор: Croft S. L., et al. 2006. Clin.Microbiol.Rev., 19, 111-126). Одной из таких мишеней являются ферменты биосинтеза полиаминов, что обусловлено необходи-мостью Put и Spd как таковых для размножения паразитов, а также и тем, что Spd входит в состав трипанотиона (Try, N1,N8-бис(глутатионил)спермидин), выполняющего многочислен-ные функции по защите паразита от неблагоприятных внешних воздействий. Одна из слож-ностей практического применения ингибиторов биосинтеза Spd для лечения протозойных инфекций состоит в том, что Spd жизненно необходим и для клеток хозяина. Тем не менее, a-дифторметилорнитин (DFMO), ингибитор ODC, используется для лечения поздних стадий сонной болезни, вызываемой Tripanosoma brucei gambiense (Burri C. C. & Brun R. 2003. Para-sitol.Res., 90, S49–S52).
Среди многочисленных ингибиторов ODC одним из наиболее эффективных является 1-аминоокси-3-аминопрпан (АРА), действующий на изолированный фермент в nM концент-рациях и специфически подавляющий рост нормальных и опухолевых клеток (Hyvoven T., et al. 1988. J. Biol. Chem., 263, ; Persson L., et al. 1989. Biochem. J., 257, 929-931; Sta-nek J., et al. 1992. J. Med. Chem., 35, ; Milovic V., et al. 2001. Biochem. Pharmacol., 61, 199-206).
 Оказалось, что АРА высокоактивен и по отношению L. donovani и P. falciparum, подав-ляя их размножение в mM концентрациях, снижая уровни Put, Spd и Try у парази-тов и был намного активнее DFMO, клас-сического ингибитора ODC (Табл. 14).
Оказалось, что АРА высокоактивен и по отношению L. donovani и P. falciparum, подав-ляя их размножение в mM концентрациях, снижая уровни Put, Spd и Try у парази-тов и был намного активнее DFMO, клас-сического ингибитора ODC (Табл. 14).
Табл. 14. APA эффективно ингибирует раз-множение P.falciparum и амастигот L.donovani и изолированную ODC этих паразитов.
Размножение и L.donovani и P.falciparum, полностью восстанавливается после добавления в среду Put/Spd, что указывает на специфичность действия ингибитора (данные не приведены).
Высокая активность АРА по отношению ODC L.donovani и P.falciparum стимулирова-ла рентгеноструктурные структурные исследования E-I комплекса. АРА правильно связыва-ется в активном центре фермента (протонированная при фи-зииологическом рН аминогруппа ингибитора выполняет якорные функции), что как и при связывании субстрата приводит к разрыву С=N-двойная связь внутреннего альдимина, образованного карбонильной группой
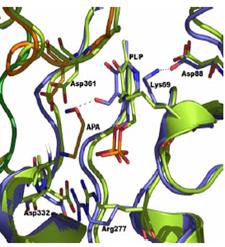
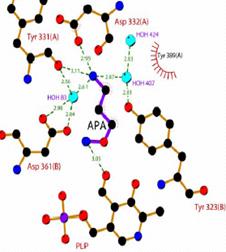 кофермента и аминогруп-пой Lys69. Пиридиновый цикл поворачивается и карбонильная группа PLP оказывается сближенной с аминоокси группой инги-битора (Рис. 28). Однако, в этом случае, в отличие от
кофермента и аминогруп-пой Lys69. Пиридиновый цикл поворачивается и карбонильная группа PLP оказывается сближенной с аминоокси группой инги-битора (Рис. 28). Однако, в этом случае, в отличие от
Рис.28. Структура комплекса ODC и APA (разрешение 1.9Å)
комплексов пиридоксаль-5’-фосфат зависимых аспартатамино-трансферазы с аминооксиук-сусной кислотой (Markovic-Housley Z., et al. 1996. Eur.J.Biochem., 236, ), трансами-назы g-аминомасляной кислоты с аминооксиуксусной кислотой (Liu W., et al. 2004. Bioche-mistry, 43, ) и 1-аминоциклопропан-1-карбоксилат синтазы с аминооксианалогом декарбоксилированного S-аденозилметионина (Capitani G., et al. 2003. Biochim.Biophys.Acta, 1647, 55-60), образование оксима фермента не наблюдалось, а карбонильная - и аминоокси-группы оказывались «замороженными» на расстоянии ~ 3Å, что соответствует длине водо-родной связи. Таким образом, показано, что в данном случае комплекс ODC–АРА кристал-лизуется в конформации, которая не реализуется в растворе, где происходит образование ок-сима фермента. Из литературы известно, что кристаллическая структура не всех комплексов ODC–DFMO соответствует строению, образующегося в растворе, ковалентного аддукта ин-гибитора с ферментом.
Успех применения специфических ингибиторов для практической терапии во многом определяется возможностью создания их активно-транспортируемых форм. Однако, такие ингибиторы ODC как DFMO и APA, весьма активные in vitro по отношению к L.donovani и P.falciparum, пассивно проникают в паразиты. Поэтому в заключительной части настоящего исследования мы попытались создать активно-транспортируемую форму APA, превратив его в GAPA, который представляет собой зарядодефицитный (рКа 6.71, см. Рис. 27) изостерный аналог агматина. Так как агматин активно проникает в животные клетки используя Put транспортер, то можно было предположить, что и GAPA сможет проникать в клетки, исполь-зуя систему транспорта Put.
Сопоставление активностей APA и GAPA по отношению к изолированной ODC (10-9 М и 6·10-6 M, соответственно) и амастиготам L.donovani (5 mM и 9 mM, соответственно) свиде-тельствует, что GAPA, скорее всего, активно проникает в паразит и, следовательно, может считаться первым активно-транспортируемым ингибитором ODC. Возможность внутрикле-точной трансформа-ции GAPA в APA (Рис. 29) определяется наличием в клетке низкоспеци-
 фичных уреидогидролаз.
фичных уреидогидролаз.
Рис. 29. Возможные механизмы действия GAPA.
Известно, что многие аргиназы спо-собны превращать канаванин в кана-лин, а некоторые из них способны даже расщеплять Agm до Put (Dabir S., et al. 2005. Int.J.Biol.Sci., 1, 114-122). Кроме того, мак-рофаги, в которых локализована L.donovani, обладают агматиназной активностью (Sastre M., et al. 1998. Biochem.J., 330, ). Проникнув в L.donovani, GAPA подобно APA вызыва-ет снижение уровня Put и Spd (Табл.15) а экзогенные Put и/или Spd полностью нейтрализуют эффекты ингибитора (данные не приведены), что свидетельствует о связи биохимической мишени GAPA с метаболизмом полиаминов.
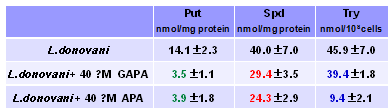 Однако GAPA, в отличие от APA, мало влияет на уровень трипанотиона в L.donovani (Табл.15). Таким образом, наблю-даемая картина, по-видимому, не
Однако GAPA, в отличие от APA, мало влияет на уровень трипанотиона в L.donovani (Табл.15). Таким образом, наблю-даемая картина, по-видимому, не
Табл. 15. Влияние GAPA и APA на уро-вень полиаминов и трипанотиона (Try) у промастигот L.donovani.
может быть описана только в терминах ингибирования ODC под действием GAPA или в ре-зультате ее расщепления до APA (Рис. 29), а взаимосвязь уровень Spd – уровень трипаноти-она у L.donovani может иметь достаточно сложный характер.
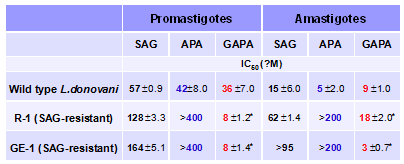 Другим принципиальным отличием GAPA от APA и DFMO является его высокая эф-фективность по отношению к формам L.donovani, резистентным к сурьмасодержащим (SAG) препаратам (Табл. 16). Следует отметить, что устойчивость к этим препаратам сопровожда-ется повышенной активность ферментов биосинтеза Spd, в первую очередь ODC, что, по-видимому, и снижает эффектив-ность АРА и DFMO.
Другим принципиальным отличием GAPA от APA и DFMO является его высокая эф-фективность по отношению к формам L.donovani, резистентным к сурьмасодержащим (SAG) препаратам (Табл. 16). Следует отметить, что устойчивость к этим препаратам сопровожда-ется повышенной активность ферментов биосинтеза Spd, в первую очередь ODC, что, по-видимому, и снижает эффектив-ность АРА и DFMO.
![]() Табл. 16. Влияние SAG, APA и GAPA на SAG-резистентные (R-1 и GE-1) клинические изоляты L.donovani.
Табл. 16. Влияние SAG, APA и GAPA на SAG-резистентные (R-1 и GE-1) клинические изоляты L.donovani.
Таким образом, нельзя исклю-чить, что GAPA имеет еще одну метаболическую мишень, отличную от ODC. Нечто подоб-ное наблюдалось ранее при сравнении эффектов DFMO и APA на рост мицелия фитопато-генного гриба Pyr.oryzae. Оба ингибитора ODC снижали содержание Put и замедляли рост мицелия, который восстанавливался при внесении в среду Put. Однако, APA, действуя, в от-личие от DFMO, еще и на ранние этапы биосинтеза меланина, вызывал обесцвечивание ми-целия, окраска которого восстанавливалась после добавления Put (, и др. 1989. Биоорган. Химия, 15, 706-709).
Превращение АРА в GAPA привело к активно проникающему в L.donovani ингибитору ODC, который, в отличие от АРА, оказался эффективен по отношению к SAG-усойчивым формам паразита. Эти приципиальные отличия в действии АРА и GAPA должны стимулировать детальное изучение механизма действия последнего, поскольку возможность существования у GAPA биологической мишени отличной от ODC, может быть уникальной для этого паразита и представлять интерес для избирательного воздействия на L.donovani.
Выводы.
1. Разработан новый подход к превращениям спермина (Spm) и спермидина (Spd) в их биологически-активные производные, заключающийся в создании системы новых С-моно-метилированных аналогов Spd, Spm и N1-Ac-Spd, для получения которых были разработаны оригинальные схемы синтезов. Ключевые соединения получены в количествах, достаточных для проведения испытаний на лабораторных животных.
2. Впервые показано, что биохимические свойства С-моно-метилированных аналогов Spm и Spd зависят от положения метильной группы. Перемещение последней карди-нально сказывается на взаимодействии аналогов с ферментами метаболизма полиами-нов, что позволило получить метаболически устойчивое производное Spd – g-MeSpd или, наоборот – индуцировать катаболическую нестабильность (b-MeSpd). Это откры-вает спектр новых возможностей для исследования клеточных функций Spm и Spd, которые легко взаимопревращаются и частично взаимозаменяемы. Найдено, что b-MeSpd, в отличие от подавляющего большинства аналогов Spd и остальных С-метилированных производных Spd, слабо индуцирует биосинтез регуляторного белка антизима, что делает b-MeSpd уникальным инструментом исследования важнейших для клетки антизим-зависимых путей поддержания гомеостаза полиаминов.
3. При помощи катаболически устойчивого a-МеSpd впервые удалось обратить последствия супериндукции спермин/спермидин-N1-ацетилтрансферазы in vitro и in vivo и предотвратить развитие острого панкреатита, вызываемого активацией катаболизма полиаминов, у крыс с регулируемой экспрессией гена SSAT.
4. Введение метильной группы в молекулы Spm и Spd приводит к возникновению хиральных центров, что открывает возможность осуществлять регуляцию биохими-ческих свойств таких аналогов полиаминов не только посредством перемещения ме-тильной группы, но и на более тонком уровне, изменяя конфигурацию хирального центра. С этой целью, были разработаны удобные методы синтеза и получены неопи-санные ранее (R)- и (S)-изомеры a-MeSpd и a-MeSpm, (R,R)-, (S,S)- и (R,S)-диастерео-меры a, a’-Me2Spm, а также (R)- и (S)-изомеры N8-Ac-a-MeSpd.
5. Впервые показано, что ферменты метаболизма полиаминов – сперминоксидаза, дезоксигипузинсинтаза и ацетилполиаминооксидаза, природные субстраты которых ахиральны, обладают скрытой стереоспецифичностью. Найден оригинальный способ регулирования стереоспецифичности ацетилполиаминооксидазы. Установлено, что система активного транспорта полиаминов по-разному узнает стереоизомеры a-мети-лированных полиаминов в качестве Spm и Spd. Способность изомеров a-МеSpd и диастереомеров a, a’-Me2Spm по-разному индуцировать +1 сдвиг рамки считывания мРНК антизима, приводящий к биосинтезу активного белка, а также продуктивный сплайсинг мРНК SSAT открывают новые возможности воздействия на ключевые регуляторные механизмы, ответственные за поддержание гомеостаза полиаминов в клетке.
6. Предложен новый подход к созданию зарядодефицитных аналогов Spm, состоящий в сокращении расстояния между аминогруппами до двух метиленовых звеньев, и син-тезированы неизвестные ранее 1,12-диамино-3,6,9-триазадодекан (SpmTrien) и его биохимически-значимые производные. Наличие триэтилентетраминового фрагмента в молекуле SpmTrien сообщает аналогу комплексообразующие свойства в отношении ионов Cu2+. Впервые показано, что введение в систему ионов меди приводит к измене-нию конденсированного состояния ДНК, вызываемого SpmTrien, что не известно для аналогов Spm.
7. На основании анализа взаимодействия SpmTrien и триэтилентераамина (Trien) с ферментами катаболизма полиаминов и эффектов, вызываемых этими аналогами в культуре клеток, показано, что SpmTrien способен выполнять многие клеточные функции Spm, тогда как Trien, изостерный Spd, не является функционально активным миметиком Spd. Установлено, что концентрация эффективно хелатирующего ионы Cu2+ SpmTrien в клетках оказывается почти в 10 раз выше, чем Trien, и что SpmTrien частично метаболизирует в клетке до Trien. Последний, благодаря образованию прочных комплексов с ионам Cu2+, в настоящее время используется в терапии нарушений миокарда, возникающих на поздних стадиях диабета II типа, а также для лечения болезни Вильсона.
8. Предложен общий способ превращения О-замещенных гидроксиламинов в ранее малодоступные алкоксигуанидины. Использовании трифлата ди-Вос-гуанидина позволяет получать разнообразные функционально-замещенные алкоксигуанидины, включая и биологически-активный 1-гуанидиноокси-3-аминопропан (GAPA) с выходами близкими к количественным.
9. Предложен способ превращения 1-аминоокси-3-аминопропана, специфического ингибитора орнитиндекарбоксилазы (скорость-определяющий фермент биосинтеза полиаминов), в GAPA – активно-транспортируемый ингибитор/проингибитор этого фермента. Показано, что GAPA эффективно ингибирует размножение Leishmania donovani, в том числе и формы резистентные к широко используемым в практической терапии лейшманиоза сурьмасодержащим препаратам.
Основное содержание работы отражено в следующих публикациях:
1. Khomutov A. R., Svetsov A. S., Vepsalainen J. J., Kritzyn A. M. "Novel acid-free cleavage of N-(2-hydroxyarylidene) protected amines". Tetrahedron Lett., 42(15), 2887-2
2. Ruiz-Chica A. J., Khomutov A. R., Medina M. A., Sanchez-Jimenez F., Ramirez F. J. "Interaction of DNA with an aminooxy analogue of spermidine - an FT-IR and FT-Raman approach". J. Mol. Structure, 565-566, 253-
3. Turchanowa L, Shvetsov A. S., Demin A. V., Khomutov A. R., Wallace H. M., Stein J., Milovic V. "Insufficiently charged isosteric analogue of spermine: interaction with polyamine uptake, and effect on Caco-2 cell growth". Biochem. Pharmacology, v. 64, No 4, p. 649-
4. Rasanen T. L., Alhonen L., Sinervirta R., Keinanen T., Herzig K-H., Suppola S., Khomutov A. R., Vepsalainen J., Janne J. "A polyamine analogue prevents acute pancreatitis and restores early liver regeneration in transgenic rats with activated polyamine catabolism". J. Biol. Chem., 277(42),
5. , “Ингибирование ферментов биосинтеза полиаминов субстратоподобными О-замещенными гидроксиламинами”. Биохимия, 67(10), 1403-1
6. , , , Хомутов А.Р. "Новый синтез α-мелилспермидина". Биоорган.химия, 30(4), 441-
7. Jarvinen A., Grigorenko N., Khomutov A. R., Hyvonen M. T., Uimari A., Vepsalainen J., Sinervirta R., Keinanen T. A., Vujcic S., Alhonen L., Porter C. W., Janne J. "Metabolic stability of alpha-methylated polyamine derivatives and their use as substitutes for the natural polyamines". J. Biol. Chem., 280(8), 6595-
8. , , Хомутов А.Р. "Новый синтез α-мелил - и α,α'-диметилспермина". Биоорган. химия, 31(2), 200-
9. , , , "Новые оксааналоги спермина". Биоорган. химия, 31(2), 206-
10. , , , "Зарядодефицитный аналог спермина с комплексообразующими свойствами" Биоорган. химия, 31,(3), 303-
11. Grigorenko N. A., Vepsalainen J., Jarvinen A., Keinanen T. A., Alhonen L., Janne J., Khomutov A. R. "Synthesis of (R)- and (S)-isomers of 1-methylspermidine" Mendeleev Commun., 15,(4), 142-
12. DasGupta R., Krause-Ihle T., Bergmann B., Muller I. B., Khomutov A. R., Muller S., Walter R. D., Luersen K. "3-Aminooxy-1-aminopropane and derivatives have an antiproliferative effect on cultured Plasmodium falciparum by decreasing intracellular polyamine concentrations". Antimicrob. Agents Chemother., 49(7), 2857-2
13. , , "Новые зарядодефицитные аналоги агматина". Биоорган. химия, 31(6), 645-
14. Jarvinen A. J., Cerrada-Gimenez M., Grigorenko N. A., Khomutov A. R., Vepsalainen J. J., Sinervirta R. M., Keinanen T. A., Alhonen L. I., Janne J. E. "a-Methyl polyamines: efficient synthesis and tolerance studies in vivo and in vitro. First evidence for dormant stereospecificity of polyamine oxidase". J. Med. Chem., 49(2), 399-
15. Jarvinen A., Keinanen T. A., Grigorenko N., Khomutov A. R., Uimari A., Vepsalainen J., Narvanen A., Alhonen L., Janne J. "Guide molecule-driven stereospecific degradation of a-methylpolyamines by polyamine oxidase". J. Biol. Chem., 281(8), 4589-4
16. Hyvonen M. T., Herzig K-H., Sinervirta R., Albrecht E., Nordback I., Sand J., Keinanen T. A., Vepsalainen J., Grigorenko N., Khomutov A. R., Kluger B., Janne J., Alhonen L. “Activated polyamine catabolism in acute pancreatitis - methylated polyamine analogues prevent trypsinogen activation and pancreatitis-associated mortality”. Am. J.Pathol., 168(1), 115-
17. , "Аминооксианалоги спермина и их моноацетильные производные." Биоорган. химия, 32(6), 643-
18. Singh S., Mukherjee A., Khomutov A. R., Persson L., Heby O., Chatterjee M., Madhubala R. “Antileishmanial effect of 3-aminooxy-1-aminopropane is due to polyamine depletion”. Antimicrob Agents Chemother. 51(2), 528-
19. Khomutov A. R., Grigorenko N. A., Skuridin S. G. “Novel approach to design isosteric charge-deficient analogue of spermine and its biochemical important derivatives”. Biochem. Soc. Trans., 35(2), 369-
20. Grillo M. A., Battaglia V., Colombatto S., Rossi C. A., Simonian A. R., Salvi M., Khomutov A., Toninello A. “Inhibition of agmatine transport in liver mitochondria by new charge-deficient agmatine analogues”. Biochem. Soc. Trans., 35(2), 401-
21. Grigorenko N. A., Khomutov A. R., Keinänen T. A., Järvinen A., Alhonen L., Jänne J., Vepsäläinen J. “Synthesis of novel optical isomers of a-methylpolyamines”. Tetrahedron, 63(10), 2257-2
22. Keinänen T. A., Järvinen A., Uimari A., Vepsäläinen J., Khomutov A. R., Grigorenko N. A., Hyvonen M. T., Cerrada-Gimenez M., Alhonen L., Jänne J. “a-Methylated polyamines as potential drugs and experimental tools in enzymology” Mini Rev. Med. Chem., 7(8), 813-
23. Dufe V. T., Ingner D., Heby O., Khomutov A. R., Persson L., Al-Karadaghi S. “A structural insight into the inhibition of human and Leishmania donovani ornithine decarboxylase by 3-aminooxy-1-aminopropane” Biochem. J., 405(2), 261-
24. Hyvonen M. T., Keinanen T. A., Cerrada-Gimenez M., Sinervirta R., Grigorenko N., Khomutov A. R., Vepsalainen J., Alhonen L., Janne J. “Role of hypusinated eukaryotic translation initiation factor 5A in polyamine depletion-induced cytostasis”. J. Biol. Chem., 282(48),
25. Simonian A., Khomutov A., Hyvonen T., Grigorenko N., Keinanen T., Vepsalainen J., Alhonen L., Janne J. " Novel CoA-polyamine conjugates for effective inhibition of spermine/spermidine-N1-acetyltransferase." Nucleosides, Nucleotides & Nucleic Acids. 26(10), 1245-1
26. Merentie M., Uimari A., Pietila M., Sinervirta R., Keinänen T. A., Vepsäläinen J., Khomutov A., Grigorenko N., Herzig K-H., Jänne J., Alhonen L. “Oxidative stress and inflammation in the pathogenesis of activated polyamine catabolism-induced acute pancreatitis”. Amino Acids, 33(2), 323-
27. Singh S., Jhingran A., Sharma A., Simonian A. R., Soininen P., Vepsalainen J., Khomutov A. R., Madhubala R. “Novel agmatine analogue, gamma-guanidinooxypropylamine (GAPA) efficiently inhibits prolifereation of Leishmania donovani by depletion of intracellular polyamine levels”. Biochem. Biophys. mun., 375(1), 168-
28. , , Умари A., Пиетила M., Серрада-Хименес M., , “Метилированные аналоги биогенных полиаминов спермина и спремидина как инструмент исследования клеточных функций полиаминов и ферментов их метаболизма“. Молекулярная биология, 43(2), 274-
29. Hyvonen M. T., Howard M. T., Anderson C. B., Grigorenko N., Khomutov A. R., Vepsalainen J., Alhonen L., Janne J., Keinanen T. A. “Divergent regulation of the key enzymes of polyamine metabolism by chiral alpha-methylated polyamine analogs”. Biochem. J., 422(2), 321-
30. Nayvelt I., Hyvönen M. T., Alhonen L., Pandya I., Thomas T., Khomutov A. R., Vepsäläinen J., Patel R., Keinänen T. A., Thomas T. J. “DNA condensation by chiral a-methylated polyamine analogues and protection of cellular DNA from oxidative damage” Biomacromolecules, 11(1), 97-
31. Weisell J., Hyvönen M. T., Vepsäläinen J., Alhonen L., Keinänen T. A., Khomutov A. R., Soininen P. ”Novel isosteric charge-deficient spermine analogue – 1,12-diamino-3,6,9-triazadodecane: Synthesis, pKa measurement and biological activity”. Amino Acids, 38(2), 501-
32. Hyvönen M. T., Sinervirta R., Keinänen T. A., Fashe T., Grigorenko N., Khomutov A. R., Vepsäläinen J., Alhonen L. “Acute pancreatitis induced by activated polyamine catabolism is associated with coagulopathy of a-methylated polyamine analogs on haemostasis” Pancreathology 10(2-3), 208-
33. Vuohelainen S., Pirinen E., Сerrada-Gimenez M., Keinanen T. A., Uimari A., Pietila M., Khomutov A. R., Janne J., Alhonen L. “Spermidine is indispensable in differentiation of 3T3-L1 fibroblasts to adipocytes” J. Cell Mol. Med., 14(6B), 1683-1
34. Weisell J., Hyvönen M. T., Häkkinen M. R., Grigorenko N. A., Pietilä M., Lampinen A., Kochetkov S. N., Alhonen L., Vepsäläinen J., Keinänen T. A., Khomutov A. R. “Synthesis and biological characterization of novel charge-deficient spermine analogs”. J. Med. Chem., 53(15), 5738-5
35. Khomutov A. R., Weisell J, Khomutov M. A., Grigorenko N. A., Simonian A. R., Häkkinen M. R., Keinänen T. A., Hyvönen M. T., Alhonen L., Kochetkov S. N., Vepsäläinen J. ”Methylated polyamines as research tools”. In: Polyamines: Methods and Protocols, Methods in Molecular Biology. (A. E.Pegg & R. A.Casero, Eds.) vol. 720, Ch.29, pp. 449-461. Springer Science+Business Media, LLC (2011).
36. Hyvönen M. T., KeinänenT. A., Khomutov M., Simonian A., Weisell J., Kochetkov S. N., Vepsäläinen J., Alhonen L., Khomutov A. R. “The use of novel C-methylated spermidine derivatives to investigate the regulation of polyamine metabolism”. J. Med. Chem., 54(13), 4611-4
37. Hyvönen M. T., Keinänen T. A., Khomutov M., Simonian A., Vepsäläinen J., Park J. H., Khomutov A. R., Alhonen L., Park M. H. “Effects of novel C-methylated spermidine analogs on cell growth via hypusination of eukaryotic translation initiation factor 5A”. Amino Acids - в печати
[*] Совместно с Prof. J. Janne, Prof. L. Alhonen, и Dr. T. Keinanen (A. I.Virtanen Institute for Molecular Sciences, University of Eeastern Finland, Kuopio, Finland).
[†] Совместно с Prof. J. Janne, Prof. L. Alhonen, Prof. J. Vepsalainen, Dr. T. Keinanen и Dr. M. Hyvonen (A. I.Virtanen Institute for Molecular Sciences, University of Eeastern Finland, Kuopio, Finland).
[‡] Совместно с Prof. J. Janne, Prof. L. Alhonen, Prof. J. Vepsalainen, Dr. T. Keinanen и Dr. M. Hyvonen (A. I.Virtanen Institute for Molecular Sciences, University of Eeastern Finland, Kuopio, Finland), а также Dr. H. M.Park (National Institute of Health, Bethesda, MD, USA).
[§] Совместно с д. б.н. , ИМБ им. В.А. Энгельгардта РАН.
[**] Совместно с Prof. R. Madhubala (J. Nehru University, New Delhi, India), Prof. L. Persson (University of Lund, Sweden), Prof. S. Al-Karadaghi (University of Lund, Sweden) и Prof. R.Walter (Institute of Tropical Medicine, Hamburg, Germany).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
