
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ü рефлексы выпрямления, направлены на поддержание естественной позы организма или ее восстановление в случае изменения. Так, например, благодаря перераспределению тонуса шейных мышц голова постоянно поддерживается в вертикальном положении.
Ø статокинетические рефлексы, направлены на сохранение позы и ориентацию в пространстве при изменении скорости движения (т. е. при действии на организм прямолинейных и угловых ускорений). Статокинетические рефлексы представляют собой резкие фазные ответы, отличные от медленных позных реакций, характерных для статических рефлексов. Они вовлекают в реакцию, как правило, почти всю мускулатуру тела, тогда как статические рефлексы – какие-то определенные группы мышц, сокращение которых обеспечивает поддержание или изменение позы. Примером статокинетических рефлексов является резкое ответное сокращение мускулатуры туловища и конечностей, возникающее при внезапной остановке или при крутом повороте автомобиля и направленное на преодоление действующих на организм ускорений. При осуществлении статокинетических рефлексов, возникающих при действии на организм угловых ускорений, немаловажное значение имеет и вовлечение в реакцию некоторых мышц глазодвигательного аппарата, ритмическое сокращение которых приводит к ритмическим движениям глазных яблок (глазной нистагм), что обеспечивает сохранение нормальной зрительной ориентации.
Вместе с тем связь вестибулярных ядер с нейронами других ядер ромбовидной ямки (как двигательными, так и парасимпатическими) обуславливает возможное вовлечение в комплекс ответных реакций организма, возникающих при возбуждении вестибулорецепторов, не только определенных скелетных мышц, но и ряда внутренних органов. Так, при действии на организм линейных или угловых ускорений (адекватных раздражителей для статокинетических вестибулярных рефлексов), наряду с перераспределением тонуса определенных скелетных мышц, участвующих в поддержании какой-то позы или ее изменении, изменяется и функциональная активность тех внутренних органов, в иннервации которых принимают участие нейроны вегетативного ядра блуждающего нерва. В частности, при длительном или чрезмерном раздражении вестибулорецепторов внутреннего уха возможно возникновение комплекса вегетативных реакций, проявляющегося в изменении сердечной деятельности, артериального давления, секреторной и моторной функции пищеварительного тракта и т. д. Причем у некоторых индивидов с высокой чувствительностью вестибулярного аппарата такие вегетативные сдвиги могут носить довольно выраженный характер и являются причиной развития симптомокомплекса, известного в клинике под названием морской болезни.
Ретикулярная формация ромбовидного мозга, наряду с образованием жизненно важных дыхательного и сосудодвигательного центров, а также со специфическим влиянием (через посредство ретикулоспинального пути) на мотонейроны спинного мозга, оказывает и определенное неспецифическое воздействие как на различные нейронные элементы спинного мозга, так вышерасположенных отделов самого головного мозга (в том числе кору больших полушарий). Так, установлено, что ретикулярная формация ромбовидного мозга оказывает на спинной мозг неспецифическое тормозное воздействие, которое затрагивает не только мотонейроны, но и первичные афферентные нервные волокна и вторичные афферентные нейроны, образующие восходящие проводящие пути к головному мозгу. Причем такое тормозное влияние ретикулярной формации на афферентные нервные элементы спинного мозга приводит к уменьшению притока чувствительной информации из спинного мозга к головному и в конечном итоге может способствовать ослаблению возбуждения в коре головного мозга и, как следствие, засыпанию. Вместе с тем на саму кору больших полушарий нейроны ретикулярной формации оказывают, как правило, неспецифическое возбуждающее влияние, способствуя поддержанию ее в активном состоянии. Следовательно, характер функционального состояния самой коры больших полушарий будет во многом зависеть от баланса тормозных и возбуждающих влияний, оказываемых ретикулярной формацией соответственно на спинной мозг и корковые нейроны.
СРЕДНИЙ МОЗГ
Средний мозг (mesencephalon) является частью ствола головного мозга, расположен между мостом и промежуточным мозгом. Нижней его границей с вентральной стороны служит верхняя горизонтальная борозда, отделяющая мост от среднего мозга, а верхней границей – зрительные тракты и сосцевидные тела (структуры промежуточного мозга). Нижней границей среднего мозга с дорсальной стороны является место выхода блокового нерва (IVп), а верхней границей – задние края таламических бугров (структур промежуточного мозга). На уровне среднего мозга из центральной нервной системы выходят две пары черепномозговых нервов – глазодвигательный нерв (IIIп) и блоковый нерв (IVп).
Средний мозг включает
Ø с вентральной стороны ножки мозга,
Ø с дорсальной стороны крышу среднего мозга (пластинку четверохолмия),
Ø в центре его находится Сильвиев водопровод (водопровод мозга) – узкий канал, соединяющий полость ромбовидного мозга (IV желудочка головного мозга) с полостью промежуточного мозга (III желудочек головного мозга).
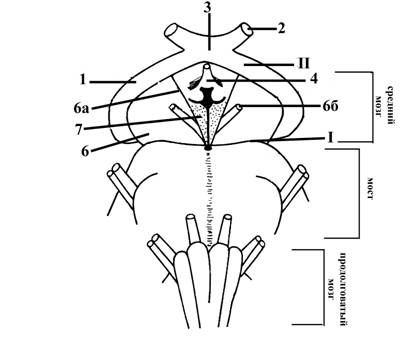
Рис. 7. Часть ствола головного мозга с вентральной стороны
I – нижняя граница среднего мозга с вентральной стороны (верхняя горизонтальная борозда)
II – верхняя граница среднего мозга с вентральной стороны (представлена зрительными трактами (1), которые являются продолжением зрительных нервов (2) после их перекреста (3) и серым бугром (4) – структурами промежуточного мозга)
6 – ножки мозга
6а – медиальная борозда ножек мозга (проходит по их медиальной поверхности), является местом выхода глазодвигательного нерва, IIIп (6б)
7 – заднее прордырявленное вещество (образует дно межножковой ямки)
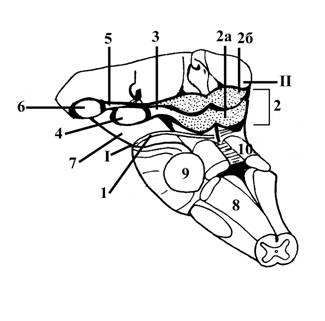
Рис. 8. Часть ствола головного мозга с дорсальной стороны
I – нижняя граница среднего мозга с дорсальной стороны – место выхода блокового нерва, IVп (1)
II – верхняя граница среднего мозга с дорсальной стороны, соответствует задним краям (подушкам) таламических бугров
2 – крыша среднего мозга – пластинка четверохолмия
2а – нижние холмики – подкорковые центры слуха, с помощью ручек нижнего холмика (3) связаны с медиальными коленчатыми телами (4) – структурами промежуточного мозга, подкорковыми центрами слуха
2б – верхние холмики – подкорковые центры зрения, с помощью ручек верхнего холмика (5) связаны с латеральными коленчатыми телами (6) – структурами промежуточного мозга, подкорковыми центрами зрения
7 – ножки мозга (вид с дорсальной стороны).
На рисунке также обозначены:
 8 – нижние мозжечковые ножки
8 – нижние мозжечковые ножки
9 – средние мозжечковые ножки
10 – верхние мозжечковые ножки
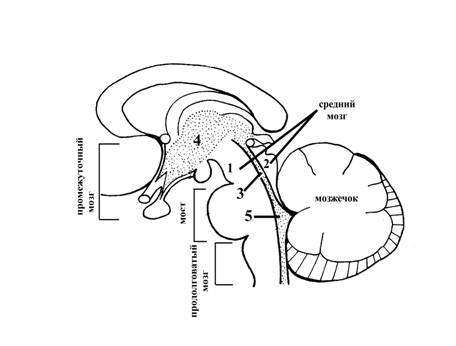
Рис. 9. Ствол головного мозга, сагиттальный разрез
1 – ножка среднего мозга
2 – крыша среднего мозга (пластинка четверохолмия)
3 – водопровод мозга (сильвиев водопровод), соединяет полость промежуточного мозга (полость III желудочка) с полостью ромбовидного мозга (полостью IV желудочка) (5)
Ножки мозга, в свою очередь, представляют собой два продольно исчерченных толстых валика, которые выходят под острым углом из моста и направляются вперед и латерально к правому и левому полушариям большого мозга. Углубление между правой и левой ножками мозга носит название межножковой ямки. Дно этой ямки является местом проникновения в ткань мозга кровеносных сосудов, в связи с чем оно изрешечено многочисленными отверстиями и составляет заднее продырявленное вещество. Вдоль медиальной поверхности ножек среднего мозга проходит медиальная или глазодвигательная борозда, которая служит местом выхода из среднего мозга глазодвигательного нерва (IIIп).
Крыша среднего мозга (пластинка четверохолмия) состоит из четырех бугорков (или холмиков), отделенных друг от друга двумя пересекающимися под прямым углом бороздами (продольной и поперечной). Поперечная борозда отделяет верхнее двухолмие от нижнего, продольная борозда – правое двухолмие от левого. Верхняя часть продольной борозды служит местом залегания шишковидной железы (эпифиза). Крыша среднего мозга характеризуется определенной локализацией серого и белого вещества в ней. Так, серое вещество (образующее ядра верхних и нижних холмиков) занимает внутреннее положение, а белое окружает его снаружи. Верхние холмики пластинки четверохолмия с помощью ручки верхних холмиков связаны с латеральными коленчатыми телами (структурами промежуточного мозга), а нижние – посредством ручки нижних холмиков – с медиальными коленчатыми телами. Ручки верхних и нижних холмиков представляют собой пучки аксонов нейронов коленчатых тел, следующих к нейронам соответствующих холмиков пластинки четверохолмия. Верхние холмики пластинки четверохолмия среднего мозга вместе с латеральными коленчатыми телами промежуточного мозга имеют отношение к обработке зрительной информации и выступают в роли подкоркового зрительного центра, тогда как нижние холмики вместе с медиальными коленчатыми телами, принимающие участие в обработке слуховой информации, – слухового подкоркового центра. Часть нейронов ядер верхнего и нижнего холмиков пластинки четверохолмия образуют нисходящий тектоспинальный путь к спинному мозгу, волокна которого еще на уровне среднего мозга перекрещиваются (перекрест Мейнерта). Тектоспинальный путь делает возможным координацию деятельности нейронов спинного мозга в зависимости от той зрительной и слуховой информации, которая поступает к соответствующим подкорковым центрам головного мозга.
Водопровод среднего мозга заполнен церебральной жидкостью и окружен узким слоем центрального серого вещества, в котором залегают ядра глазодвигательного (на уровне верхних холмиков) и блокового (на уровне нижних холмиков) нервов, а также одно из ядер ретикулярной формации среднего мозга (промежуточное ядро). Блоковый нерв (IVп) имеет только одно двигательное ядро, обеспечивающее эфферентную иннервацию верхней косой мышцы глазного яблока. Глазодвигательный нерв (IIIп) характеризуется наличием комплекса эфферентных ядер, среди которых выделяют:
Ø двигательные ядра, обеспечивающие эфферентную иннервацию некоторых мышц глазного яблока (мышца, поднимающая верхнее веко, прямые верхняя, нижняя, медиальная мышцы, нижняя косая мышца).
Ø 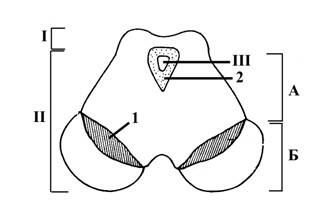 вегетативные парасимпарические ядра, обеспечивающие парасимпатическую иннервацию мышцы – сфинктера зрачка (добавочное ядро или ядро Якубовича) и мышцы цилиарного (ресничного) тела (ядро Перля).
вегетативные парасимпарические ядра, обеспечивающие парасимпатическую иннервацию мышцы – сфинктера зрачка (добавочное ядро или ядро Якубовича) и мышцы цилиарного (ресничного) тела (ядро Перля).
Рис. 10. Схема внутреннего строения среднего мозга на фронтальном сечении
Средний мозг включает:
I – крышу среднего мозга (пластинку четверохолмия)
II – ножки мозга
III – Сильвиев водопровод
Ножки мозга(II) черной субстанцией Земмеринга (1) делятся на основание (Б) и покрышку (А).
Вокруг Сильвиевого водопровода (III) находится окружающий его слой серого вещества (2), в нижней вентральной части которого на уровне нижнего двухолмия залегает ядро блокового нерва (IVп), а на уровне верхнего двухолмия–ядра глазодвигательного нерва (IIIп).
В толще ножек мозга поперечно на всем их протяжении проходит черная субстанция Земмеринга, разделяющая ножки среднего мозга на основание и покрышку. Черная субстанция представлена дофаминергическими нейронами, содержащими пигмент меланин, который обуславливает их темную окраску. Эти нейроны двусторонне связаны с базальными ганглиями большого мозга, посылают свои аксоны к ретикулярной формации ствола головного мозга, красному ядру среднего мозга; участвуют в сложной координации движений, в регуляции эмоционального поведения.
В основании ножек мозга, в свою очередь, проходят нисходящие проводящие пути преимущественно пирамидной системы (кортикоспинальный, кортикооливарный, кортикоретикулярный, кортикомостовой, кортиковестибулярный и некоторые другие), представляющие собой пучки аксонов пирамидных клеток коры больших полушарий, связывающие кору большого мозга с различными отделами ствола головного мозга и со спинным мозгом.
В покрышке ножек среднего мозга залегают красное ядро, ретикулярная формация, проходят волокна медиальной и латеральной петель. Красное ядро имеет удлиненную форму и простирается от уровня нижних холмиков до таламусов, продолжаясь в них. Нейроны этого ядра получают возбуждающие входы от коры головного мозга, черной субстанции, мозжечка (шаровидного, пробковидного и зубчатого его ядер). Аксоны же нейронов красного ядра образуют нисходящий руброспинальный тракт, который на уровне среднего мозга перекрещивается, образуя вентральный перекрест покрышки (перекрест Фореля). Данное ядро имеет отношение к сложной координации движений, оказывает стимулирующее влияние на тонус мотонейронов мышц – сгибателей.
Медиальная и латеральная петли, проходящие в покрышке ножек среднего мозга, представляют собой пучки нервных волокон, проводящих определенную чувствительную информацию к таламусу. В частности, волокна медиальной петли имеют отношение к проведению проприоцептивной, тактильной, болевой и температурной информации к таламусу, тогда как волокна латеральной петли – слуховой информации в направлении нижних холмиков пластинки четверохолмия и таламических бугров.
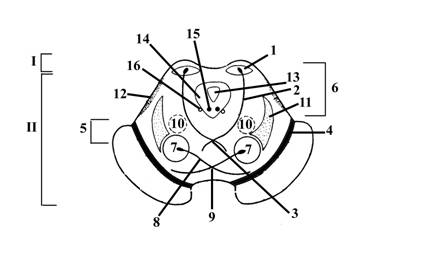
Рис. 11. Схема внутреннего строения среднего мозга на уровне нижнего двухолмия (место залегания ядер блокового нерва)
I – пластинка четверохолмия (крыша среднего мозга); показано нижнее двухолмие, где находятся ядра – подкорковые центры слуха (1).
От ядер верхнего и нижнего двухолмия начинается нисходящий тектоспинальный тракт (2), волокна которого на уровне среднего мозга перекрещиваются, образуя дорсальный перекрест покрышки (перекрест Мейнерта).
II – ножка среднего мозга
4 – черная субстанция Земмеринга, разделяет ножки мозга на основание (5) и покрышку (6).
В основании (5) проходят нисходящие проводящие пути (кортикомостовой, кортикоретикулярный. кортикооливарный, кортикоспинальный и т. д.).
В покрышке ножек мозга залегают:
7 – красное ядро; аксоны нейронов этого ядра образуют нисходящий руброспинальный тракт (8), который на уровне среднего мозга перекрещивается (вентральный перекрест покрышки – перекрест Фореля – 9)
10 – ретикулярная формация – продолжение ретикулярной формации ромбовидного мозга.
11 – медиальная петля (совокупность нервных волокон, несущих проприоцептивную, тактильную, болевую и температурную информацию к таламусу)
12 – латеральная петля (совокупность нервных волокон – аксонов нейронов слуховых ядер или ядер трапециевидного тела, несущих слуховую информацию к ядрам нижнего двухолмия); имеется в толще среднего мозга только на уровне нижних холмиков, так как волокна, ее составляющие, переключаются на нейроны ядер нижнего двухолмия – подкорковые центры слуха
13 – водопровод мозга (Сильвиев водопровод), вокруг которого находится небольшой слой серого вещества (14). В вентральной части последнего на уровне нижнего двухолмия залегает ядро блокового нерва, IVп (15) – двигательное, обеспечивает эфферентную иннервацию верхней косой мышцы глазного яблока. Аксоны же нейронов этого ядра, образующие блоковый нерв, выходят со стороны латеральной части среднего мозга, на границе его с мостом. На уровне верхнего двухолмия в вентральной части серого вещества, окружающего Сильвиев водопровод, залегают ядра глазодвигательного нерва, IIIп. Аксоны нейронов двигательных и вегетативных парасимпатических ядер глазодвигательного нерва, образующие этот нерв, выходят из головного мозга через медиальную борозду ножек среднего мозга.
16 – среднемозговое ядро тройничного нерва, чувствительное, начинается на уровне моста и продолжается в средний мозг.
Функции среднего мозга
Функции среднего мозга во многом определяются деятельностью входящих в его состав ядер. Так, вегетативные ядра глазодвигательного нерва вместе с подкорковыми зрительными центрами (верхние холмики пластинки четверохолмия) обеспечивают осуществление
Ø аккомодационного рефлекса (состоит в сокращении ресничной мышцы глазного яблока, сопровождающемся округлением хрусталика (увеличением радиуса его кривизны, а следовательно, и преломляющей способности) при рассматривании близко расположенных от глаз предметов);
Ø некоторых зрачковых рефлексов (например, сужение зрачков в ответ на увеличение интенсивности освещения сетчатки глазного яблока или на дефокусировку изображения на сетчатке, возникающую по причине перевода взгляда с дальних на ближние для глаз предметы).
Двигательные ядра глазодвигательного нерва принимают участие в осуществлении ряда глазодвигательных рефлексов, в частности, рефлекторного сведения (конвергенции) зрительных осей обоих глаз на рассматриваемом объекте.
Ядра верхних и нижних холмиков пластинки четверохолмия (подкорковые зрительные и слуховые центры среднего мозга) обеспечивают осуществление ориентировочных и сторожевых рефлексов, которые возникают в ответ на раздражение зрительных и слуховых рецепторов и протекают неосознанно (возникают еще до момента восприятия животным организмом зрительной или слуховой информации). Эти рефлексы могут осуществляться даже у животных с разрушенными большими полушариями при условии целостности среднего мозга. Зрительные и слуховые ориентировочные рефлексы проявляются в резком повороте глаз, головы, а возможно и всего туловища в направлении неожиданно возникших вспышки света или звукового сигнала. Сторожевые рефлексы, подобно ориентировочным, также возникают в ответ на неожиданные световые или звуковые сигналы, действующие на организм, как правило, с большой интенсивностью, и проявляются в повышении тонуса мышц-сгибателей, что способствует бегству или нападению животного. Сторожевые рефлексы носят, как правило, защитный характер и позволяют человеку или животному быстро реагировать на неожиданные зрительные или слуховые раздражители.
Черная субстанция среднего мозга имеет отношение к
Ø координированию сложных актов глотания и жевания, хотя рефлекторные дуги этих рефлексов замыкаются на уровне ромбовидного мозга;
Ø координации тонких движений пальцев рук;
Ø регуляции эмоционального поведения.
Причем немаловажное значение в регулирующем влиянии черной субстанции на сложные двигательные акты имеет связь ее нейронов с красным ядром среднего мозга и ретикулярной формацией ствола головного мозга, принимающих участие в образовании нисходящих рубро - и ретикулоспинального трактов, которые собственно и проводят эфферентную информацию к мотонейронам спинного мозга (эфферентным нейронам многих двигательных рефлексов). Нарушение функционального состояния черной субстанции среднего мозга является одной из причин возникновения болезни Паркинсона, проявляющейся в нарушении тонких движений пальцев рук, развитии мышечной ригидности и появлении тремора.
Красное ядро и ретикулярная формация среднего мозга принимают участие в регуляции сложных двигательных актов организма, оказывая стимулирующее влияние через посредство рубро - и ретикулоспинальных трактов преимущественно на мотонейроны, иннервирующие мышцы-сгибатели. Перерезка ствола головного мозга ниже среднего мозга сопровождается развитием у животного децеребрационной ригидности, проявляющейся в значительном повышении тонуса мышц-разгибателей на фоне резкого ослабления тонуса мышц-сгибателей. Развитие данного состояния связано с нарушением целостности рубро-, ретикуло - и кортикоспинальных трактов, оказывающих стимулирующее влияние на мотонейроны мышц-сгибателей, в результате чего начинают преобладать вестибулоспинальные влияния, повышающие активность мотонейронов мышц-разгибателей.
Наряду с выше описанной рефлекторной деятельностью среднего мозга, он выполняет и важную проводниковую функцию. Последняя обусловлена проведением через средний мозг как чувствительной информации от тактильных, болевых и температурных рецепторов кожи, проприорецепторов опорно-двигательного аппарата, висцерорецепторов внутренних органов, вестибулярных рецепторов (по волокнам медиальной петли), слуховых рецепторов (по волокнам латеральной петли) к выше расположенным отделам головного мозга, так и проведении эфферентной информации от коры больших полушарий к нейронам ствола головного мозга или спинного мозга.
МОЗЖЕЧОК
Мозжечок или малый мозг (cerebellum) располагается над стволовой частью головного мозга (прилежит сверху к продолговатому мозгу и мосту), заложен в задней черепной ямке. Сверху над мозжечком нависают затылочные доли полушарий большого мозга, которые отделены от мозжечка поперечной щелью большого мозга.
Мозжечок по своему внешнему строению напоминает большой (конечный мозг), но, являясь более древней частью центральной нервной системы, устроен значительно проще. В мозжечке выделяют два полушария (левое и правое) и соединяющую их непарную часть – червь. Полушария мозжечка характеризуются определенной топографией серого и белого вещества. В частности, серое вещество полушарий мозжечка занимает периферическое положение, образуя кору мозжечка, а белое – локализовано внутри полушарий. В толще белого вещества полушарий мозжечка, в свою очередь, находятся скопления серого вещества, образующие ядра мозжечка (аналоги базальных ганглиев большого мозга). Кора мозжечка в отличие от таковой больших полушарий является трехслойной (тогда как кора большого мозга шестислойная).
Поверхность мозжечка не является гладкой: она изрезана многочисленными щелями и бороздами. Самой глубокой является горизонтальная щель, разделяющая мозжечок на верхнюю и нижнюю поверхности. Более мелкие щели мозжечка, проходя поперечно и параллельно друг другу, не прерываясь через полушария и червь, отделяют друг от друга листки (извилины) мозжечка. Группы извилин, отделенные более глубокими бороздами, образуют дольки мозжечка. Причем каждой дольке червя соответствуют две (правая и левая) определенные дольки полушарий. Более изолированной и генетически старой долькой полушарий является клочок (flocculus), который с помощью длинной ножки клочка соединяется с соответствующей долькой червя, называемой узелком (nodulus).
Ядра мозжечка, образованные скоплениями серого вещества, заложенными внутри полушарий, являются парными и представлены следующими разновидностями:
Ø ядро шатра (занимает самое медиальной положение в полушариях)
Ø шаровидное ядро
Ø пробковидное ядро
Ø зубчатое ядро (занимает самое латеральное положение в полушариях, является самым крупным и филогенетически молодым ядром).
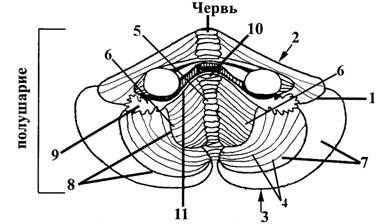
Рис. 12. Схема строения мозжечка (вид снизу и спереди)
1 – горизонтальная щель мозжечка, разделяющая его на верхнюю (2) и нижнюю (3) поверхности
4 – щели мозжечка; проходят, не прерываясь, через полушария и червь и разделяют их на дольки; следовательно, каждой дольке червя (5)соответствуют две дольки полушарий (6)
6 – дольки полушарий мозжечка, более мелкими щелями разделены на извилины или листки. Совокупность долек объединяется в более крупные доли (7), которые отделены друг от друга более глубокими щелями (8)
9 – изолированная долька полушария (клочок, flocculus)
10 – долька червя (узелок, nodulus), с которой соединяются с помощью флокулонодулярных ножек (11) изолированные дольки полушарий (клочки).
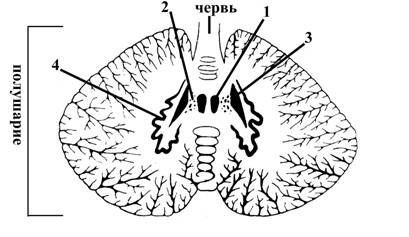
Рис. 13. Схема ядер мозжечка на горизонтальном разрезе
1 – ядро Шатра, связано с вестибулярными ядрами моста, ведает равновесием тела
 2 – шаровидное ядро осуществляют координацию движений
2 – шаровидное ядро осуществляют координацию движений
3 – пробковидное ядро туловища
4 – зубчатое ядро (осуществляет координацию движений конечностей)
Шаровидное, пробковидное и зубчатое ядра связаны с ретикулярной формацией ствола головного мозга, красным ядром среднего мозга, нижнеоливарными ядрами продолговатого мозга (посылают к этим структурам ствола головного мозга свои аксоны)
В мозжечке с филогенетической точки зрения выделяют три отдела:
Ø архиоцеребеллюм (древний мозжечок), к которому относятся ядро шатра вместе с клочком (flocculus) и узелком (nodulus),
Ø палеоцеребеллюм (старый мозжечок), включающий червь вместе с шаровидным и пробковидным ядрами,
Ø неоцеребеллюм (новый мозжечок), представленный полушариями мозжечка (кроме клочка) вместе с зубчатым ядром.
Мозжечок связан с различными отделами ствола головного мозга с помощью трех пар мозжечковых ножек – нижних средних и верхних. Мозжечковые ножки представляют собой совокупность нервных волокон, следующих из определенных отделов ствола головного мозга в кору мозжечка, или из мозжечка к определенным структурам мозгового ствола. В частности, нижние мозжечковые ножки связывают мозжечок с продолговатым мозгом, средние – с мостом, а верхние – со средним мозгом.
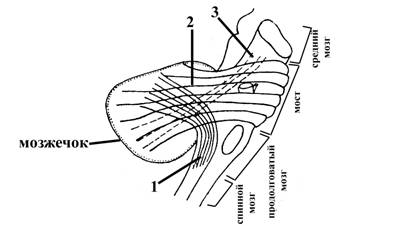
Рис. 14. Схематическое изображение направление хода ножек мозжечка (вид сбоку)
1 – нижние ножки мозжечка (пучки нервных волокон, связывающие мозжечок с продолговатым мозгом)
2 – средние ножки мозжечка (пучки нервных волокон. связывающие мозжечок с мостом)
3 – верхние ножки мозжечка (пучки нервных волокон, связывающие средний мозг с мозжечком).
В составе нижних мозжечковых ножек проходят:
ü нервные волокна дорсального спиномозжечкового тракта;
ü часть аксонов нейронов ядер Голя и Бурдаха;
ü аксоны нейронов вестибулярных ядер;
ü аксоны нейронов нижнеоливарных ядер;
ü аксоны нейронов ядра шатра, направляющиеся к вестибулярным ядрам;
ü аксоны нейронов шаровидного, пробковидного и зубчатого ядер, направляющиеся к ретикулярной формации продолговатого мозга.
В составе средних мозжечковых ножек проходят:
ü нервные волокна – аксоны нейронов собственных ядер моста
В составе верхних мозжечковых ножек проходят:
ü нервные волокна вентрального спиномозжечкового тракта;
ü нервные волокна шаровидного, пробковидного и зубчатого ядер, связывающие их с нейронами красного ядра и таламуса.
Функциональные взаимоотношения коры и ядер мозжечка состоят в следующем. Все афферентные входы в мозжечок (следующие в составе его ножек из ствола головного мозга) направляются в кору мозжечка, где вступают в синаптический контакт с определенными нейронами коры. Кора мозжечка, в свою очередь, образует единственный эфферентный выход, представленный аксонами клеток Пуркинье (грушевидных нейронов). Аксоны грушевидных нейронов коры мозжечка направляются либо к его ядрам, либо, не переключаясь на нейронах мозжечковых ядер, напрямую следуют к определенным структурам ствола головного мозга (в составе соответствующих мозжечковых ножек). Аксоны нейронов мозжечковых ядер выходят из мозжечка и также следуют к определенным структурам мозгового ствола. Таким образом, первоначальная обработка всей входящей в мозжечок информации осуществляется в его коре, после чего может продолжаться в его ядрах, а затем уже эфферентная информация проводится от мозжечка к определенным структурам ствола мозга (например, к красному ядру, ретикулярной формации и некоторым другим). Вместе с тем возможен и вариант обработки чувствительной информации, поступающей к мозжечку, только на уровне его коры, после чего эфферентная информация из коры мозжечка напрямую (минуя его ядра) проводится к определенным нейронам ствола мозга.
Структурная организация коры мозжечка
Кора мозжечка характеризуется исключительной упорядоченностью составляющих ее нервных элементов. В частности, в ней выделяют три слоя:
Ø молекулярный (самый поверхностный)
Ø ганглиозный (занимает срединное положение)
Ø зернистый (самый внутренний, прилежит к белому веществу полушарий мозжечка).
В молекулярном слое залегают тела корзинчатых и звездчатых нейронов, а также аксоны вставочных нейронов (тела которых находятся в зернистом слое коры) и дендриты грушевидных нейронов (тела которых залегают в ганглиозном слое коры). Ганглиозный слой коры мозжечка образован скоплением грушевидных клеток (клеток Пуркинье) – самых крупных клеток коры мозжечка, на которых размещается до 20000 синапсов, образуемых другими нервными элементами (как аксонами нейронов собственно коры мозжечка, так и нервными волокнами, приходящими в мозжечок из ствола головного мозга). В зернистом слое коры мозжечка, в свою очередь, залегают тела вставочных нейронов (или зернистых клеток) и клеток Гольджи. В функциональном отношении только вставочные нейроны коры мозжечка являются возбуждающими, тогда как все остальные клетки (грушевидные нейроны, звездчатые, корзинчатые клетки и клетки Гольджи) – тормозными. Вставочных нейронов примерно в 2300 раз больше, чем грушевидных нейронов.
Единственный эфферентный выход из коры мозжечка образуют аксоны грушевидных нейронов (клеток Пуркинье), тогда как все остальные нейроны коры передают информацию в конечном итоге на грушевидные клетки (непосредственно, вступая с ними в синаптический контакт, или опосредованно через какие-то другие нейроны коры мозжечка). Так, звездчатые, корзинчатые и вставочные нейроны образуют синапсы непосредственно на нейронах Пуркинье. При этом вставочные нейроны образуют возбуждающие синапсы преимущественно на дендритах грушевидных нейронов, а корзинчатые и звездчатые клетки – тормозные синапсы на телах грушевидных нейронов (такие тормозные синапсы непосредственно на телах обеспечивают довольно эффективное торможение грушевидных нейронов). Клетки Гольджи непосредственно в контакт с нейронами Пуркинье не вступают, а образуют тормозные синапсы на телах и дендритах вставочных нейронов, ограничивая их активность. Вместе с тем, как было сказано выше, вставочные нейроны представляют собой единственный тип нейронов коры мозжечка, который оказывает возбуждающее действие на клетки Пуркинье. Таким образом, клетки Гольджи, хотя сами и не оказывают тормозного влияния на грушевидные нейроны, но тормозят активность вставочных нейронов, способных возбуждать клетки Пуркинье. Следовательно, корзинчатые и звездчатые нейроны могут непосредственно, а клетки Гольджи опосредованно (через вставочные нейроны) тормозить активность клеток Пуркинье, аксоны которых образуют единственный эфферентный выход из коры мозжечка. Вставочные нейроны, напротив, могут оказывать возбуждающее влияние на грушевидные клетки.
Клетки Пуркинье, в свою очередь, являются тормозными нейронами и их аксоны образуют тормозные синапсы на иннервируемых структурах, будь-то нейроны ядер самого мозжечка или непосредственно нейроны ядер ствола головного мозга.
Чувствительная информация в кору мозжечка поступает по двум афферентным входам:
Ø лазающим волокнам, представляющим собой аксоны нейронов нижнеоливарного ядра продолговатого мозга (промежуточного ядра равновесия)
Ø мшистым (моховидным) волокнам, представляющим собой аксоны вестибулярных ядер ромбовидного мозга, ядер Голля и Бурдаха продолговатого мозга, собственных ядер моста, вентрального и дорсального спиномозжечковых трактов.
Оба афферентных входа в кору мозжечка являются возбуждающими, но отличаются друг от друга не только своим происхождением, но и степенью разветвленности, а также тем, с какими типами нейронов коры мозжечка они взаимодействуют. Так, лазающие волокна (аксоны нейронов нижнеоливарных ядер продолговатого мозга), практически не разветвляясь, вступают в синаптический контакт непосредственно с клетками Пуркинье, на которые оказывают возбуждающее действие. Мшистые волокна, многократно разветвляясь (в 20 и более раз), вступают в синаптический контакт со всеми остальными нейронами коры мозжечка (за исключением самих клеток Пуркинье) – звездчатыми, корзинчатыми, вставочными нейронами и клетками Гольджи, на которые также оказывают возбуждающее действие. Вместе с тем, поскольку почти все нейроны коры мозжечка, за исключением вставочных нейронов, являются тормозными, то возбуждающие влияния мшистых волокон на иннервируемые ими нейроны коры мозжечка будут в большинстве своем (за исключением влияний, передаваемых через вставочные нейроны) приводить к торможению активности клеток Пуркинье.
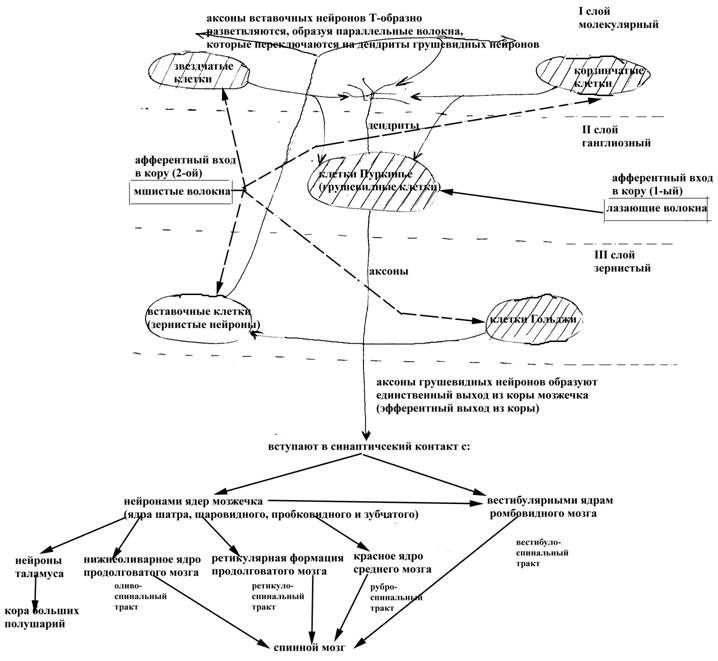 |
Рис. 15. Схема структурной организации коры мозжечка (заштрихованы тормозные нейроны, не заштрихованы возбуждающие нейроны)
Таким образом, в кору мозжечка поступает информация от рецепторов опорно-двигательного аппарата (по волокнам дорсального и вентрального спиномозжечковых трактов, аксонам нейронов ядер Голля и Бурдаха), вестибулярных рецепторов (по волокнам вестибулярных ядер ромбовидного мозга) и некоторых других структур нервной системы, а единственный эфферентный выход из коры мозжечка, который является тормозным, направляется либо к нейронам ядер самого мозжечка, либо непосредственно к структурам ствола головного мозга (в частности, к вестибулярным ядрам ромбовидного мозга). Аксоны же нейронов ядер мозжечка также следуют к определенным структурам мозгового ствола, таким как нижнеоливарное ядро продолговатого мозга, ретикулярная формация продолговатого мозга, красное ядро среднего мозга, таламические бугры, вестибулярные ядра ромбовидного мозга и некоторым другим. Аксоны нейронов большинства из этих структур мозгового ствола, в свою очередь, принимают участие в образовании нисходящих проводящих путей к спинному мозгу (рубро-, ретикуло-, оливо-, вестибулоспинальные тракты), в котором заложены собственно тела мотонейронов, иннервирующих почти всю мускулатуру туловища и конечностей. Таким путем мозжечок через посредство различных структур ствола головного мозга может обеспечивать, наряду с другими отделами центральной нервной системы, регуляцию двигательных актов организма. Наконец, функциональное состояние нейронов коры мозжечка может отчасти регулироваться и корой больших полушарий. Так, аксоны пирамидных нейронов Беца коры больших полушарий, образующие нисходящие пирамидные тракты, частично переключаются на нейроны вестибулярных, нижнеоливарных ядер и собственных ядер моста, которые, в свою очередь, посылают свои аксоны в кору мозжечка. Таким образом, через посредство названных ядер ствола головного мозга кора больших полушарий может регулировать функциональное состояние мозжечка.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
