
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таким образом, различные нейроны коры специализируются на выполнении определенных функций. Так, афферентые входы, поступающие в кору головного мозга (преимущественно аксоны нейронов таламических ядер и базальных ганглиев), вступают в синаптический контакт в основном с клетками III (наружный пирамидный) и IV (внутренний зернистый) слоев. Аксоны же нейронов III и IV слоев коры переключаются либо непосредственно на нейроны V слоя (внутренний пирамидный), образующего единственный эфферентный выход из коры, либо вступают в синаптический контакт с нейронами II (наружный зернистый) и VI (мультиформный) слоев, выполняющих функцию вставочного звена в коре больших полушарий и передающих информацию в конечном итоге на нейроны внутреннего пирамидного слоя (слой V).
На основании особенностей клеточного состава и функционального значения кору больших полушарий разделяют на ряд участков, называемых корковыми полями. Широким международным признанием пользуется цитоархитектоническая карта Бродмана, описавшего у человека 52 клеточных поля. Наряду со сложной шестислойной клеточной структурой коры (цитоархитектоникой), имеется и сложная структура ее нервных волокон (миелоархитектоника), одни из которых обеспечивают связи по горизонтали в пределах какого-то слоя коры (горизонтальные волокна), а другие – связи по вертикали, как между различными участками коры, так и с другими отделами центральной нервной системы (радиальные волокна).
Функциональное значение различных областей коры больших полушарий
С функциональной точки зрения в коре больших полушарий можно выделить три типа зон, специализированных на выполнении определенных функций:
Ø сенсорные
Ø ассоциативные
Ø моторные.
Сенсорные зоны представляют собой области, в которые поступает информация от различных рецептивных полей организма (корковые представительства анализаторов) и где осуществляется высший ее анализ. Причем каждая сенсорная зона специализируется на обработке информации от рецептивного поля какого-то одного или нескольких анализаторов (зрительного или слухового или кожного, висцероцептивного, проприоцептивного и т. д.). Удаление или генерализованное повреждение этих зон сопровождается потерей человеком определенных видов чувствительности.
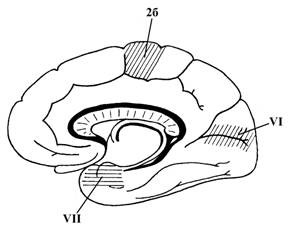
В коре головного мозга человека выделяют две сенсорные зоны – I и II соматосенсорные зоны, специализированные на обработке соматосенсорной информации (от кожных рецепторов, рецепторов опорно-двигательного аппарата, внутренних органов и сосудов), зрительную, слуховую, вкусовую и обонятельную сенсорные зоны (см. рис.13).
I соматосенсорная зона расположена в постцентральной извилине и верхней теменной дольке, а II соматосенсорная зона – под центральной бороздой и распространяется на верхний край сильвиевой борозды. Обе эти зоны получают чувствительную информацию от рецепторов кожи, опорно-двигательного аппарата и внутренних органов противоположной стороны тела через посредство заднего вентрального ядра таламуса. Причем наибольшую площадь в этих зонах занимает корковое представительство рецепторов кисти рук, голосового аппарата и лица, а наименьшую – туловища, бедра и голени. Площадь коркового представительства рецептивного поля определенной части тела определяется количеством нейронов, участвующих в восприятии и анализе афферентной информации от этого поля (чем количество корковых нейронов больше, тем более дифференцированным оказывается анализ раздражений).
Зрительная сенсорная зона расположена в медиальной поверхности затылочных долей полушарий, по обе стороны от шпорной борозды, а слуховая сенсорная зона – в латеральной поверхности височной доли больших полушарий, на уровне первой поперечной извилины Гешля. Вкусовая сенсорная зона расположена в височной доле, вблизи от сильвиевой и циркулярной борозды, а корковое представительство обонятельного анализатора соответствует медиальной поверхности височной доли, в области крючка.
Моторные зоны коры (см. рис.13) по своей функции являются эфферентными и принимают участие в регуляции и осуществлении различных двигательных актов организма. Они расположены в области предцентральной извилины и околоцентральной дольки (медиальная поверхность полушарий) и характеризуются средоточием большого количества гигантских пирамидных клеток Беца в V слое коры. Аксоны этих пирамидных нейронов Беца в составе пирамидного тракта направляются к нейронам базальных ганглиев, ядер мозгового ствола и спинного мозга, которые прямо или опосредованно принимают участие в иннервации различных групп скелетных мышц. Следовательно, связь моторной зоны коры с собственно мотонейронами, заложенными в стволе головного мозга и спинном мозге, осуществляется через посредство пирамидных трактов (кортико-ядерных и кортикоспинальных). В связи с тем, что большая часть волокон пирамидных трактов образует перекрест на уровне мозгового ствола или границы продолговатого мозга со спинным, моторные области каждого из полушарий имеют отношение к регуляции деятельности мышц противоположной стороны тела. Вместе с тем деятельность мышц туловища, гортани и глотки регулируется как правым, так и левым полушариями. Электрическое раздражение определенных участков моторных зон коры сопровождается сокращением определенных скелетных мышц или даже отдельных их двигательных единиц, а генерализованное повреждение этих зон влечет за собой частичный или полный паралич мускулатуры противоположной стороны тела. Наряду с выработкой в моторной зоне коры эфферентной программы действий, передаваемой в дальнейшем к определенным группам мотонейронов, здесь также происходит и анализ проприоцептивных (кинетических) раздражений, возникающих в работающих мышцах. В связи с этим моторная зона коры очень тесно функционально связана с I соматосенсорной зоной. Подобно соматосенсорной зоне, в моторной зоне наибольшую площадь занимает корковое нейронное представительство мускулатуры кистей рук, лица, губ, языка, а наименьшую – туловища и нижних конечностей. В зависимости от площади коркового моторного представительства будет иметь место та или иная степень точности и тонкости управления движениями данной части тела (наибольшей она является для кистей рук, языка, мышц лица).
Ассоциативные зоны располагаются по краю сенсорных зон (во все стороны от них на 1-5см). Подобно сенсорным зонам коры, ассоциативные зоны имеют отношение к обработке чувствительной информации, поступающей от различных рецептивных полей, но в отличие от сенсорных зон, специализированных на обработке определенного типа чувствительной информации, принимают участие в сложном сочетанном анализе информации, поступающей от рецептивных полей различных анализаторов. Удаление ассоциативных зон не влечет за собой потерю определенного вида чувствительности, но сопровождается нарушением способности правильно интерпретировать значение действующего раздражителя; причем в большинстве своем подобные нарушения являются временными, благодаря широким компенсаторным возможностям других неповрежденных ассоциативных зон. Так, разрушение у человека 18-го и 19-го полей по К. Бродману, являющихся вторичной зрительной зоной, не приводит к возникновению слепоты, но у человека нарушается способность оценивать виденное (в частности, он не понимает значения слов при чтении).
А Б
Рис.13. Схема расположения корковых концов анализаторов
А – вид на верхнелатеральную поверхность полушария
Б – вид на медиальную поверхность полушария
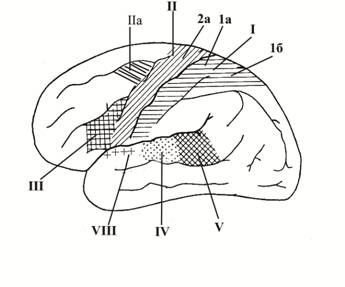
I – I соматосенсорная зона, в которой происходит анализ чувствительной информации, поступающей от рецепторов кожи (тактильных, болевых, температурных), опорно-двигательного аппарата и внутренних органов (т. е. эта зона представляет собой корковый конец кожного, проприоцептивного и висцероцептивного анализаторов); находится в постцентральной извилине (1а, соответствует полям 1, 2 и 3 по К. Бродману) и верхней теменной дольке (1б, соответствует полям 5 и 7 по К. Бродману). Проводящие чувствительные пути, следующие от рецепторов кожи, опорно-двигательного аппарата, внутренних органов, на уровне спинного мозга или ствола головного мозга перекрещиваются, вследствие чего I соматосенсорные зоны каждого полушария получают информацию от рецепторов противоположной части тела. Кроме того, в постцентральной извилине и верхней теменной дольке рецептивные поля различных участков тела спроецированы таким образом, что наиболее высоко (ближе к средней линии головы) расположены корковые представительства рецептивных полей нижних отделов туловища и нижних конечностей, а наиболее низко (ближе к сильвиевой борозде) – корковые представительства рецептивных полей верхних участков тела, верхних конечностей и головы (иными словами корковое представительство рецептивных полей кожи и опорно-двигательного аппарата тела человека является "перевернутым верх ногами"). В верхней теменной дольке находится ядро кожного анализатора, которому присуща функция узнавания предметов на ощупь (стереогнозия).
II – моторная зона, располагается в предцентральной извилине (2а, соответствует полям 4 и 6 по К. Бродману) и околоцентральной дольке (медиальная поверхность полушарий, 2б). Причем в верхних участках предцентральной извилины и в парацентральной дольке расположены клетки, функционально связанные с мотонейронами, иннервирующими мышцы самых нижних отделов туловища и нижних конечностей. В нижней части предцентральной извилины, наоборот, находятся двигательные центры, регулирующие деятельность мышц шеи, головы и лица (т. е. подобно чувствительным проекциям различных областей тела в I соматосенсорной зоне, в моторной зоне коры двигательные центры нижней и верхней части тела также являются спроецированными "верх ногами"). Неподалеку от моторной зоны (в средней лобной извилине, заднем ее отделе, поле 8 по К. Бродману) находится двигательная зона письменной речи (IIа), контролирующая деятельность мышц, принимающих участие в произвольных движениях руки при написании букв и других знаков. Эта двигательная зона тесно связана с теми участками моторной зоны предцентральной извилины, которые контролируют движение руки (соответствующей полю 40 по К. Бродману) и сочетанный поворот головы и глаз в противоположную сторону (средняя лобная извилина, поле 8). Повреждение двигательной зоны письменной речи не приводит к нарушению всех видов движений, а сопровождается лишь утратой способности производить рукой точные и тонкие движения при написании букв, знаков и слов (аграфия). В области предцентральной извилины, соответствующей полю 40, залегает также двигательное ядро, осуществляющее синтез всех целенаправленных сложных комбинированных движений (одностороннее, у правшей находится в левом полушарии, а у левшей – в правом). Деятельность этого ядра совершенствуется в течение жизни (вследствие формирования временных связей между его нейронами в результате практической деятельности и накопления опыта). Поражение этого ядра не вызывает паралича мышц, а лишь приводит к потере способности производить сложные координированные движения (апраксии).
III – речедвигательная зона (моторная зона), находится в задней части нижней лобной извилины – двигательный центр Брокка; односторонний (у "правшей" находится в левом полушарии, у "левшей" – в правом). Функциональна связана с теми отделами предцентальной извилины, которые анализируют движения, производимые при сокращении мышц головы и шеи (в том числе, и движения всех мышц (губ, щек, языка, гортани), принимающих участие в акте образования устной речи). Повреждение участка речедвигательной зоны (соответствующего полю 44 по К. Бродману) приводит к двигательной афазии, т. е. утрате способности произносить слова. Причем такая афазия не связана с потерей способности к сокращению мышц, участвующих в речеобразовании, более того у человека сохраняется способность к произношению звуков и к пению. Повреждение же соседнего поля 45 сопровождается вокальной амузией – неспособностью к составлению и воспроизведению музыкальных фраз и аграмматизмом – утратой способности к составлению осмысленных предложений из отдельных слов.
IV – ядро слухового анализатора, находится в средней части верхней височной извилины (извилины Гешля) на поверхности, обращенной к островку (поля 41, 42, 45).
V – ядро слухового анализатора устной речи (центр Вернике), располагается в задней части верхней височной извилины, в глубине латеральной борозды; функция этого ядра заключается в анализе как слышимой человеком чужой речи, так и своей собственной. Поражение этой области коры не нарушает слухового восприятия звуков вообще, но утрачивается способность понимать слова, речь (словесная глухота или сенсорная афазия). В средней же трети верхней височной извилины находится ядро слухового анализатора, поражение которого сопровождается наступлением музыкальной глухоты, при которой музыкальные фразы воспринимаются как бессмысленный набор различных шумов.
VI – ядро зрительного анализатора, располагается в затылочной доле (медиальной ее поверхности) по краям от шпорной борозды (поля 17, 18, 19). Двусторонне поражение ядра зрительного анализатора человека приводит к полной слепоте. Поражение же только какой-то части этого ядра, например, поля 18 сопровождается потерей зрительной памяти при сохраненном зрении. Вблизи ядра зрительного анализатора в нижней теменной дольке находится функционально связанное с ним ядро зрительного анализатора письменной речи, поражение которого приводит к утрате способности воспринимать написанный текст и читать его (алексия)
VII – ядро обонятельного анализатора, находится в области медиальной поверхности височной доли, нижней ее части (в крючке и отчасти в области гиппокампа)
VII – ядро вкусового анализатора, находится в височной доле, вблизи латеральной борозды и самых нижних отделов пост центральной извилины (поле 43).
Электрическая активность коры (понятие об электроэнцефалограмме)
Электрическую активность коры больших полушарий (т. н. электроэнцефалограмму), отражающую уровень ее функциональной активности, можно зарегистрировать, используя электроэнцефалограф. При этом существует два способа отведения биопотениалов, связанных с активностью нейронов коры от поверхности головы:
Ø биполярное отведение, оба электрода активны, поскольку прикладываются к поверхности кожных покровов черепа в области проекций определенных участков коры (под ними постоянно изменяется разность потенциалов в процессе регистрации), в данном случае регистрируется изменение разности потенциалов между этими электродами);
Ø монополярное отведение, один электрод – активный – прикладывается к поверхности черепа в области проекции коры, а другой – пассивный, потенциал под которым в процессе регистрации практически не изменяется – к коже носа или к мочке уха; в данном случае регистрируется изменение потенциала под активным электродом.
Амплитуда потенциалов, отводимых от кожных покровов черепа человека, колеблется в пределах от 5-10 до 200-300мкВ, а частота от 0,5 до 70 колебаний и более в секунду. Причем чем более частым является ритм, тем он менее высоко амплитудный и наоборот. Происхождение волн электроэнцефалограммы большинство специалистов связывают не с суммацией потенциалов действия множества асинхронно работающих нейронов, а с суммацией постсинаптических потенциалов, синхронно или асинхронно возникающих в различных участках коры, в зависимости от характера прихода к ним афферентной импульсации. Так, при синхронном возбуждении нервных клеток коры (что, как правило, имеет место при ограничении поступления афферентной информации при закрывании глаз, пребывании в тихом и затемненном помещении, во время сна и наркоза) в результате суммации постсинаптических потенциалов на отводящих электродах регистрируются медленные, но высокоамплитудные волны. В период активного бодрствования приток афферентной информации от различных рецептивных полей в кору резко возрастает, причем постсинаптические потенциалы в различных нейронах возникают асинхронно (в связи с резким усилением аффернтного притока) и на электроэнцефалограмме регистрируются частые, но низкоамплитудные ритмы.
По частоте, амплитуде и физиологическим характеристикам электрических колебаний на электроэнцефалограмме различают четыре основных типа волн:
Ø a-ритм – ритмические колебания потенциала почти синусоидной формы (амплитуда 50мкв, частота 8-13 колебаний в секунду), возникает при условии физического и умственного покоя, в положении лежа или сидя с расслабленной мускулатурой и закрытыми глазами, при отсутствии внешних раздражений. При таком состоянии организма данный ритм наиболее выражен в затылочной и теменной долях коры
Ø b-ритм (амплитуда 20-25мкВ, частота – свыше 13 в секунду), наблюдается при переходе из покоящегося состояния в активное, когда возрастает приток афферентной импульсации к коре. Так, например, при нанесении световых раздражителей, умственной работе, эмоциональном возбуждении он появляется в затылочных долях, при физической активности – в пред - и постцентральной извилинах
Ø q-ритм (амплитуда 100-150мкВ, частота – 4-8 в секунду) и d-ритм (амплитуда 250-300мкВ, частота – 0,5-3,5 в секунду), наблюдаются при резком уменьшении притока чувствительной информации к коре, например, во время сна, а также при некоторых патологических состояниях (при гипоксии, глубоком наркозе, патологических состояниях в самой коре).
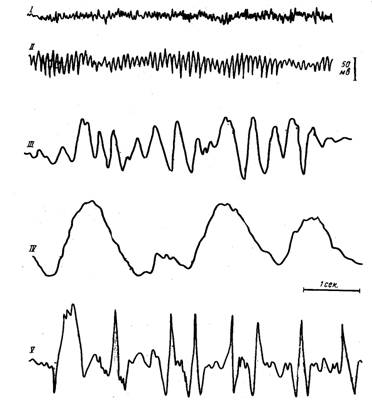
Рис. 14. Основные ритмы электроэнцефалограммы (схема)
I – b-ритм
II – a-ритм
III – q-ритм
IV – d-ритм
V – судорожные разряды
Морфо-функциональная характеристика базальных ганглиев
Базальные ганглии это скопления тел нервных клеток (серое вещество) в толще белого вещества больших полушарий головного мозга. Они являются продолжением стволовых структур головного мозга (поэтому их еще называют подкорковыми образованиями).
К базальным ганглиям относят:
Ø миндалевидное тело, залегает в белом веществе переднемедиального отдела височной доли полушария, является подкорковым обонятельным центром, имеет отношение к регуляции деятельности многих внутренних органов (пищеварительной системы, матки и некоторых др.), играет важное значение в процессах памяти
Ø ограда (имеет вид тонкой вертикально расположенной пластинки серого вещества), залегает между корой островковой доли и чечевицеобразным ядром полосатого тела в белом веществе теменной доли; физиологическая роль ее не ясна
Ø полосатое тело (на фронтальном и поперечном разрезе имеет вид чередующихся полос серого и белого вещества, благодаря чему и получило такое название), располагается сбоку от таламуса, включает следующие структуры:
ü хвостатое ядро, в котором выделяют головку (располагается в лобной доле полушарий, представляет собой утолщенную переднюю часть хвостатого ядра), тело (залегает в области дна центральной части бокового желудочка, в белом веществе лобной и теменной долей) и хвост (истонченный участок хвостатого ядра, который достигает миндалевидного тела, лежащего в височной доле)
ü чечевицеобразное ядро, по форме напоминает чечевичное зерно, залегает латеральнее от таламуса и хвостатого ядра, но медиальнее ограды, преимущественно в теменной доле больших полушарий; на фронтальном разрезе больших полушарий оно имеет треугольную форму, причем, вершина треугольника обращена в медиальную, а выпуклое основание – в латеральную сторону. Две параллельные, расположенные почти в сагиттальной плоскости прослойки белого вещества делят чечевицеобразное ядро на три части: скорлупа (занимает самое латеральное положение), латеральная и медиальная пластинки бледного шара.
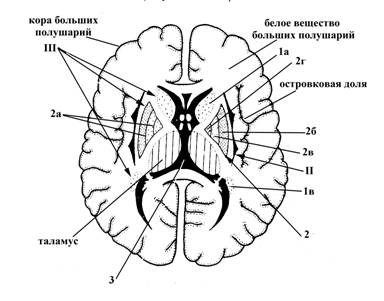
А Б
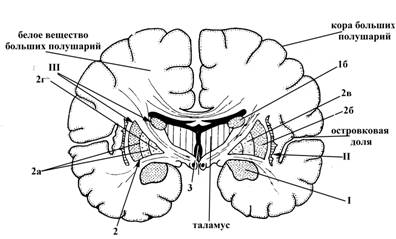
Рис.15. Полушарие большого мозга на горизонтальном (А) и фронтальном (Б) разрезе
I – миндалевидное тело, залегает в белом веществе переднемедиального отдела височной доли полушария
II – ограда (имеет вид тонкой вертикально расположенной пластинки серого вещества), залегает между корой островковой доли и чечевицеобразным ядром полосатого тела (III) в белом веществе теменной доли
III – полосатое тело (на фронтальном и поперечном разрезе имеет вид чередующихся полос серого и белого вещества), располагается сбоку от таламуса, включает следующие структуры:
1 – хвостатое ядро, в котором выделяют головку (1а) (располагается в лобной доле полушарий, представляет собой утолщенную переднюю часть хвостатого ядра), тело (1б) (залегает в области дна центральной части бокового желудочка, в белом веществе лобной и теменной долей) и хвост (1в) (истонченный участок хвостатого ядра, который достигает миндалевидного тела (I), лежащего в височной доле)
2 – чечевицеобразное ядро, по форме напоминает чечевичное зерно, залегает латеральнее от таламуса и хвостатого ядра (III), но медиальнее ограды (II), преимущественно в теменной доле больших прлушарий; на фронтальном разрезе больших полушарий оно имеет треугольную форму, причем, вершина треугольника обращена в медиальную, а выпуклое основание – в латеральную сторону. Две параллельные, расположенные почти в сагиттальной плоскости прослойки белого вещества (2а) делят чечевицеобразное ядро на три части: скорлупа (2б) (занимает самое латеральное положение), латеральная (2в) и медиальная (2г) пластинки бледного шара.
На рисунке 15 также обозначено: 3 – полость III желудочка головного мозга (полость промежуточного мозга).
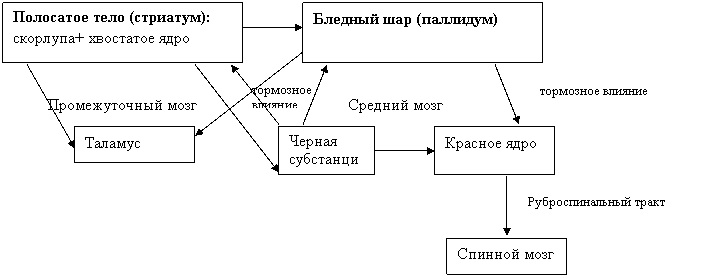
В настоящее время под названием полосатое тело (striatum) объединяют только скорлупу и хвостатое ядро – филогенетически более новые образования. Бледный шар (его медиальная и латеральная пластинки) выделяется под названием pallidum, так как его строение и развитие отличаются от других частей полосатого тела, и он является филогенетически более старым образованием. В своей совокупности striatum и pallidum составляют стриопадллидарную систему.
Стриопаллидарная система получает
Ø афферентные входы из неспецифических ядер таламуса, черной субстанции (оказывает преимущественно тормозное влияние на нейроны бледного шара и полосатого тела),
Ø эфферентные выходы из коры больших полушарий (направляются преимущественно из сенсомоторной зоны коры, причем нервные волокна корковых нейронов образуют синапсы в основном на нейронах полосатого тела, на которые оказывают возбуждающее влияние).
Полосатое тело и бледный шар посылают свои проекции (аксоны своих нейронов) к коре больших полушарий (диффузно к различным ее участкам), таламусу, черной субстанции. Кроме того, стриатум направляет часть своих нервных волокон к паллидуму и черной субстанции, а паллидум – к красному ядру среднего мозга (оказывает на нейроны красного ядра тормозное влияние). Благодаря описанным двусторонним связям стриопаллидарной системы с корой большого мозга и таламусом, с одной стороны, и черной субстанцией и красным ядром (связь только односторонняя) среднего мозга, с другой, она имеет отношение к сложной координации движений и поэтому считается составным компонентом экстрапирамидной системы. В частности, в электрофизиологических исследованиях показано, что электрическое раздражение хвостатого ядра, подобно непосредственной аппликации дофамина на его нейроны, вызывает стереотипные движения головы и дрожательные движения верхних конечностей. Вещества же, тормозящие синтез дофамина или блокирующие дофаминвые рецепторы, устраняют двигательные эффекты в ответ на электрическое раздражение хвостатого ядра. Главным же источником дофаминергических нейронов, аксоны которых образуют синапсы на клетках стриопаллидарной системы, является черная субстанция среднего мозга. Патологические ее изменения ведут к уменьшению содержания дофамина в стриатуме и нарушению его деятельности, что проявляется в первую очередь в расстройствах тонкой моторики пальцев рук (болезнь Паркинсона). Двусторонние же перерезки у животных путей, идущих из черной субстанции в стриатум, вызывают отказ от еды и питья, неподвижность, отсутствие ответов на раздражение из внешнего мира.
Наряду с отмеченным имеются данные, согласно которым полосатое тело играет роль в процессах запоминания двигательных программ, а его раздражение может приводить к нарушению обучения и памяти. Установлено также, что стриопаллидарная система оказывает тормозное влияние на различные проявления двигательной активности и на эмоциональные компоненты двигательного поведения, в частности, на агрессивные реакции. Кроме того, согласно некоторым экспериментальным данным, в полосатом теле находятся высшие вегетативные координационные центры, регулирующие обмен веществ, теплообразование, тепловыделение, сосудистые реакции. Наконец, существует предположение о том, что в полосатом теле находятся центры, интегрирующие безусловно рефлекторные двигательные и вегетативные реакции в единый целостный акт поведения, причем немаловажную роль в деятельности этих центров имеет связь стриатума с гипоталамусом.
Бледный шар вместе с таламусом и гипоталамусом у млекопитающих животных и, в том числе человека, принимает участие в осуществлении сложных безусловных рефлексов – оборонительных, ориентировочных, пищевых, половых. Кроме того, через бледный шар проходят рефлекторные дуги, от которых зависят вспомогательные движения, сопровождающие всякий сложный двигательный акт. При поражениях паллидума движения становятся угловатыми, однообразными, происходит обеднение двигательных актов вспомогательными движениями, наблюдается также мимическая неподвижность лица (маскообразное лицо).
 |
 |
Понятие о лимбической системе
Под лимбической системой понимают совокупность структур головного мозга (лимбическую область среднего мозга, гипоталамус, миндалевидное ядро больших полушарий, а также древнюю и старую кору), принимающих участие в регуляции вегетативных функций, и обеспечивающих своей регулирующей деятельностью поддержание гомеостаза, самосохранение индивида и сохранение вида вообще. Так, древняя и старая кора больших полушарий, наряду с обработкой обонятельной информации
Ø получает и афферентную информацию от многих других рецептивных полей организма (благодаря своим связям с теменной, зрительной, слуховой и другими областями коры), а следовательно, играет важную роль в процессах обработки афферентной информации и эфферентного синтеза
Ø ведает реакциями настораживания и внимания,
Ø гиппокамп, будучи функционально связан с ретикулярной формацией и миндалевидным ядром, принимает непосредственное участие в формировании эмоциональных реакций
Ø принимает участие в регуляции вегетативных функций, осуществлении инстинктивного поведения (пищевого, полового, оборонительного).
Миндалевидное ядро больших полушарий имеет отношение к регуляции деятельности многих внутренних органов (пищеварительного тракта, мочевого пузыря, матки и др.). Так, при его электрическом раздражении у животных наблюдается облизывание, жевание, глотание, саливация, изменение желудочной секреции и моторики кишечника. Регулирующее влияние старой коры и миндалевидного ядра на вегетативные функции осуществляется под контролем гипоталамуса (высшего вегетативного центра). В связи с этим к лимбической системе наряду со старой и древней корой, а также базальными ганглиями, относят и гипоталамус, а также ретикулярную формацию среднего мозга.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
