
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Функциональное значение мозжечка
Мозжечок, наряду с корой больших полушарий, играет важную роль в регуляции равновесия тела и осуществлении многих сложных двигательных программ. Кроме того, благодаря связи мозжечка с ретикулярной формацией ствола головного мозга он имеет отношение и к регуляции вегетативных функций.
Электрическое раздражение различных участков мозжечка вызывает изменение электрической активности нейронов определенных участков коры больших полушарий, ядер мозгового ствола и ретикулярной формации головного мозга. При достаточно сильных электрических раздражениях поверхности мозжечка или отдельных его ядер возникают движения глаз, головы и конечностей, которые отличаются от таковых, возникающих при раздражении моторных зон коры больших полушарий, тем, что являются медленными, имеют тонический характер (являются плавными) и сохраняются долгое время.
Удаление или поражение мозжечка вызывает расстройства статических и статокинетических рефлексов, а также произвольных двигательных реакций организма. Отмеченный факт указывает на то, что влияние мозжечка, с одной стороны, распространяется на тонические рефлексы положения и выпрямления и статокинетические рефлексы, осуществляемые соответствующими центрами ромбовидного мозга, а с другой – на моторную кору и связанные с ней центры, ведающие произвольными движениями организма.
Мозжечковые расстройства у человека выражаются в том, что он при стоянии с открытыми глазами сильно шатается, а при закрывании глаз падает, ходит зигзагами; движения становятся не координированными. В целом при различных поражениях мозжечка у человека и животных возникает целый комплекс двигательных расстройств, который может иметь следующие проявления:
Ø асинергия – нарушение взаимодействия между нервными центрами различных мышц (так, например, делая шаг, больной выносит ногу, но при этом не сгибает вперед туловище, что приводит к падению), разновидностью асинергии является адиадохокинез– нарушение возможности производить быстрые чередующиеся движения антагонистическими группами мышц (например, сгибать и разгибать несколько раз подряд руку);
Ø дезэквилибрация – различные нарушения равновесия, проявляющиеся либо расстройствами статических рефлексов (например, животное не может принимать и поддерживать определенные позы), либо нарушением каких-то произвольных движений;
Ø атония – потеря тонуса определенных мышц, и в более легких случаях – дистония (нарушения регуляции мышечного тонуса)
Ø астазия – нарушение способности мышц к слитному тетаническому сокращению, в результате чего сокращение многих мышц носит не плавный, а резкий характер и голова, туловище и конечности больного непрерывно дрожат и качаются;
Ø астения – легкая утомляемость при работе скелетных мышц, возникающая вследствие того, что движения производятся неэкономично, при участии большого количества мышц, а следовательно, многие двигательные акты организма требуют большей затраты энергии, чем в норме;
Ø атаксия – недостаточная координация движений и расстройства силы, величины и скорости направления движений. Атаксия проявляется в резко выраженных нарушениях походки, которая становится шаткой; движения рук и ног при ходьбе неловкие, слишком сильные и размашистые.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг (diencephalon) является самой проксимальной (верхней) частью ствола головного мозга. Нижней его границей с дорсальной стороны служит горизонтальная борозда, отделяющая верхние холмики пластинки четверохолмия (структуры среднего мозга) от таламических бугров, а с вентральной стороны – передний край заднего продырявленного вещества и зрительные тракты; верхней границей промежуточного мозга с вентральной стороны выступает передняя поверхность зрительного перекреста. В промежуточном мозге выделяют:
Ø таламическую область (с дорсальной стороны)
Ø гипоталамическую область (с вентральной стороны).
Полостью промежуточного мозга является III желудочек головного мозга. В состав промежуточного мозга входят две железы внутренней секреции – гипофиз (анатомически и функционально связан с гипоталамусом) и эпифиз (структурно связан с таламусом).
Белое вещество промежуточного мозга представлено нервными волокнами, входящими в состав восходящих и нисходящих проводящих путей, следующих соответственно в направлении больших полушарий и нижележащих отделов мозгового ствола или спинного мозга. Серое вещество промежуточного мозга составляют ядра, которые по функции могут быть чувствительными, нейросекреторными, либо относиться к ретикулярной формации или вегетативным центрам.
Морфо-функциональная характеристика таламуса (таламической области)
Структурная организация таламуса
Таламическая область промежуточного мозга занимает дорсальное положение, представлена:
Ø собственно таламическими буграми (зрительными буграми), составляющими задний таламус,
Ø 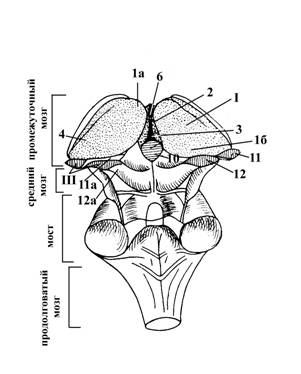 метаталамусом (заталамической областью), к которому относятся латеральные и медиальные коленчатые тела
метаталамусом (заталамической областью), к которому относятся латеральные и медиальные коленчатые тела
Ø эпиталамусом (надталамической областью), включающим эпифиз и структуры, которые обеспечивают его связь с таламическими буграми (поводки эпифиза, треугольники поводков, спайка поводков).
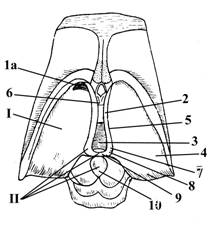
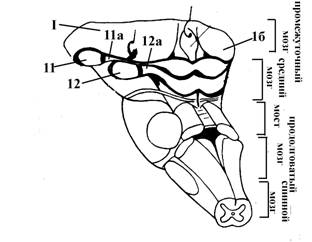
Рис.1.Ствол головного мозга Рис.2.Схема строения тала - Рис. 3. Ствол головного
с дорсальной стороны мического (зрительного) мозга (вид сзади и сбоку)
мозга
I – таламус (зрительный бугор) – подкорковый центр всех видов чувствительности за исключением обонятельной. Его передний конец сужен и заканчивается передним бугорком (1а), а задний конец утолщен и называется подушкой (1б).
2 – медиальная поверхность зрительного бугра, принимает участие в образовании латеральных (боковых) стенок III желудочка (3).
4 – верхняя поверхность таламуса, принимает участие в образовании дна боковых желудочков головного мозга (полостей большого или конечного мозга)
5 – мозговая полоска зрительного бугра (или таламуса), отделяет медиальную его поверхность (2) от верхней (4).
6 – межбугорное сращение (медиальные поверхности левого и правого таламуса в передней своей части соединены друг с другом)
II – эпиталамус (надталамическая область)
Мозговая полоска (5) кзади срастается с поводками эпифиза (8). В месте сращения мозговых полосок с поводками эпифиза образуются треугольные расширения – треугольники поводков (7). Задние отделы поводков срастаются между собой, образуя спайку поводков (9), к которой прикрепляется эпифиз (10).
Следовательно, эпиталамус (надбугорная область) включает:
o треугольники поводков (7),
o поводки эпифиза (8),
o спайку поводков (9),
o эпифиз (шишковидное тело) – (10).
III – метаталамус (заталамическая область). Представлен латеральными (11) и медиальными (12) коленчатыми телами – парными образованиями продолговато-овальной формы, соединяющимися с крышей среднего мозга (пластинкой четверохолмия) с помощью ручек верхнего (11а) и нижнего (12а) холмиков соответственно.
Зрительные бугры (образуют собственно таламус) имеют овоидную форму, расположены по обеим сторонам III желудочка, принимая участие в образовании латеральных его стенок; верхние же их поверхности формируют дно боковых желудочков головного мозга (полостей больших полушарий). В переднем отделе таламический бугор суживается и заканчивается передним бугорком, а задний его конец – расширен и носит название таламической подушки. Серое вещество таламических бугров образует до 40 ядер, разделенных тонкими прослойками белого вещества. По функциональному принципу все таламические ядра являются чувствительными (т. е. имеют отношение к обработке чувствительной информации поступающей от различных рецептивных полей организма по соответствующим проводящим путям) и посылают свои проекции к большому или конечному мозгу, на уровне коры которого осуществляется окончательная обработка всей чувствительной информации. В зависимости от характера связи таламических ядер с большими полушариями их классифицируют на две группы:
Ø специфические, которые посылают свои аксоны непосредственно к коре головного мозга (причем к строго к определенным ее участкам), аксоны нейронов этих ядер в большинстве своем не имеют коллатералей; раздражение в эксперименте специфических ядер вызывает изменение электрической активности только в ограниченных участках коры больших полушарий;
Ø неспецифические, представляющие собой продолжение ретикулярной формации ствола головного мозга, их проекции направляются к нейронам базальных ганглиев, посылающим свои аксоны диффузно к различным участкам коры (т. е. аксоны нейронов неспецифических ядер непосредственно в контакт с нейронами коры не вступают и в отличие от аксонов специфических ядер образуют множество коллатералей). Раздражение неспецифических ядер в эксперименте оказывает влияние на электрическую активность в широких областях коры больших полушарий, вызывая т. н. «реакцию активации коры».
Все ядра таламуса обязательно получают эфферентные проекции из коры головного мозга, т. е. связь их с корой больших полушарий двусторонняя.
По топографическому принципу ядра таламуса подразделяются на следующие группы (см. рис.4):
1 – передняя группа ядер (залегает в области бугорков таламусов). Представляет собой неспецифические ядра (т. е. ядра ретикулярной формации).
2 – ядра средней линии (или интраламинарные ядра, совокупность ядер, залегающих в толще боковых стенок III желудочка и в области межталамического сращения) Представляют собой ядра ретикулярной формации, имеющие отношение к регуляции (через базальные ганглии) активности нейронов черной субстанции среднего мозга (черная субстанция, в свою очередь, ведает активностью нейронов красного ядра среднего мозга). Следовательно, ядра средней линии имеют отношение к сложной координации двигательных актов и поэтому относятся к экстрапирамидной системе.
3 – срединные ядра. Представляют собой ядра ретикулярной формации, имеющие отношение к регуляции (через базальные ганглии, кору головного мозга и гипоталамус) вегетативных функций.
4 – латеральные ядра, специфические ядра таламуса, к нейронам которых поступает афферентная информация от рецепторов кожи (болевых и температурных – по волокнам латерального спинно-таламического тракта, тактильных – по волокнам вентрального спинно-таламического тракта), проприорецепторов опорно-двигательного аппарата (по волокнам проприоцептивных проводящих путей коркового направления – аксонам нейронов ядер Голя и Бурдаха продолговатого мозга). Иными словами, к нейронам латеральных ядер таламуса поступает афферентная информация по нервным волокнам медиальной петли. Эти ядра посылают афферентную информацию непосредственно к коре, причем, к строго определенным ее участкам. Так, тактильная, болевая и температурная информация проводится к нейронам постцентральной извилины, а проприоцептивная – к нейронам пост центральной и отчасти предцентральной извилины.
 5 – задняя группа ядер. Представляет собой специфические ядра таламуса; включает ядра подушек таламусов (зрительные ядра, получают афферентные входы по нервным волокнам зрительных трактов и посылают свои проекции к нейронам затылочной доли коры больших полушарий, по обе стороны от шпорной борозды – корковому концу зрительного анализатора), ядра латеральных (подкорковые центры зрения) и медиальных (подкорковые центры слуха) коленчатых тел.
5 – задняя группа ядер. Представляет собой специфические ядра таламуса; включает ядра подушек таламусов (зрительные ядра, получают афферентные входы по нервным волокнам зрительных трактов и посылают свои проекции к нейронам затылочной доли коры больших полушарий, по обе стороны от шпорной борозды – корковому концу зрительного анализатора), ядра латеральных (подкорковые центры зрения) и медиальных (подкорковые центры слуха) коленчатых тел.
Рис.4. Схематическое изображение локализации групп ядер в таламических буграх
Метаталамус (заталамическая область) занимает в таламусе латеральное положение. Представлен латеральными и медиальными коленчатыми телами – парными образованиями продолговато-овальной формы, соединенными с крышей среднего мозга (пластинкой четверохолмия) с помощью ручек верхнего и нижнего холмиков соответственно.
Латеральные коленчатые тела находятся у нижнелатеральной поверхности таламусов, сбоку от их подушек, служат подкорковыми центрами зрения. С помощью ручки верхнего холмика (пучка аксонов нейронов самого тела) каждое латеральное коленчатое тело соединено с верхним холмиком пластинки четверохолмия среднего мозга. Таким образом, ручки верхних холмиков являются эфферентными выходами латеральных коленчатых тел. С латеральными коленчатыми телами вступают в синаптическую связь часть нервных волокон зрительного тракта одноименной стороны (представляют собой афферентные входы в латеральные коленчатые тела). Часть нервных волокон латерального коленчатого тела направляется к коре головного мозга (затылочная доля, по обе стороны от шпорной борозды – корковый центр зрительного анализатора).
Медиальные коленчатые тела располагаются несколько медиальнее и кзади от латеральных, под подушками таламусов, представляют собой подкорковый центр слуха, с помощью ручек нижних холмиков соединены с нижними холмиками пластинки четверохолмия среднего мозга. Ручки нижних холмиков образованы аксонами нейронов медиальных коленчатых тел и служат их эфферентными выходами. С медиальными коленчатыми телами вступают в синаптическую связь часть нервных волокон латеральной петли (афферентные входы в медиальные коленчатые тела). Часть нервных волокон медиального коленчатого тела направляется непосредственно к коре головного мозга (височная доля, область между I и II извилинами Гешля – корковый слуховой центр).
Эпиталамус (надталамическая область) включает шишковидное тело (эпифиз) и те структуры, которые обеспечивают его прикрепление к медиальной поверхности таламических бугров (треугольники поводков, поводки эпифиза, спайка поводков).
Функциональное значение таламуса
Таламус является коллектором всех чувствительных путей (за исключением обонятельных), идущих к большим полушариям, а следовательно, на уровне таламуса осуществляется подкорковый анализ различной сенсорной информации, в связи с чем его образно называют «чувствилищем мозга». Причем тот факт, что большинство афферентных сигналов на пути к коре больших полушарий переключаются на ядрах таламуса имеет важное значение, которое заключается в том, что кора больших полушарий через посредство своих нисходящих тормозных проекций к специфическим ядрам таламуса подавляет слабые возбуждающие влияния, поступающие к таламусу от различных рецепторов. В результате такого нисходящего тормозного влияния коры больших полушарий на определенные таламические структуры обеспечивается выделение наиболее важной информации, которая и проводится в направлении коры больших полушарий.
Наряду с тем, что таламус является воротами, через которые в кору проходит почти вся чувствительная информация, и принимает участие в начальной (подкорковой) ее обработке, допускается и определенная его самостоятельная роль (т. е. без участия коры больших полушарий) в формировании у человека некоторых ощущений. Так, ряд нейрофизиологов считают таламус высшим болевым центром в связи с тем, что раздражение определенных его структур (без участия коры больших полушарий) может приводит к возникновению чувства боли у человека.
Неспецифические ядра таламуса, составляющие в своей совокупности его ретикулярную формацию, посылают свои многочисленные диффузные проекции через базальные ганглии больших полушарий к нейронам коры и оказывают неспецифическое возбуждающее влияние на кору больших полушарий, поддерживая определенный ритм ее активности, необходимый для деятельного состояния. Причем активацию неспецифических ядер таламуса наиболее эффективно вызывают болевые сигналы, поступающие от различных областей тела и внутренних органов. Следовательно, система неспецифических ядер таламуса осуществляет контроль ритмической активности коры больших полушарий и выполняет функции внутриталамической интегрирующей системы.
Наконец, некоторые специфические ядра таламуса способны оказывать регулирующее воздействие на подкорковые структуры, имеющие отношение к осуществлению ряда сложных рефлексов. Таким образом, таламус может играть важную роль в качестве надсегментарного центра рефлекторной деятельности. В частности, у животных с удаленными большими полушариями сохраняются некоторые сложные двигательные рефлексы (связанные с обеспечением локомоции, глотания, жевания, сосания), тогда как после перерезки ц. н.с. ниже таламуса эти функции нарушаются.
Морфо-функциональная характеристика гипоталамуса (гипоталамической области)
Структурная организация гипоталамуса
Гипоталамическая область занимает вентральное положение в промежуточном мозге, принимая участие в формировании дна III желудочка головного мозга. К гипоталамусу относятся зрительный перекрест, зрительный тракт, сосцевидные тела, серый бугор с воронкой и гипофизом, а также стенка дна III желудочка головного мозга, в которой заложено до 30 гипоталамических ядер.
Зрительный перекрест формируется в результате частичного перехода волокон зрительных нервов (II пара) на противоположную сторону. После зрительного перекреста следуют зрительные тракты, в составе каждого их которых проходят волокна, как левого, так и правого зрительных нервов. Зрительный тракт, в свою очередь, делится на три ветви, одна из которых направляется к подушке таламических бугров, вторая – к латеральным коленчатым телам и третья – к верхним холмикам пластинки четверохолмия. Таким образом, зрительный тракт обеспечивает проведение чувствительной информации, поступающей от сетчатки глазного яблока, к подкорковым зрительным центрам.
Кзади от зрительного перекреста находится серый бугор, позади которого лежат сосцевидные тела, а по бокам – зрительные тракты. Книзу серый бугор переходит в воронку, связанную с гипофизом.
Сосцевидные тела расположены между серым бугром спереди и задним продырявленным веществом сзади. Они имеют вид небольших (d≈0,5см) сферических образований белого цвета. Белое вещество занимает в них периферическое, а серое – центральное положение, формируя ядра сосцевидных тел. В сосцевидных телах заканчиваются столбы свода, несущие к ним обонятельную информацию из конечного мозга. Следовательно, они представляют собой подкорковые обонятельные центры.
Ядра гипоталамуса по топографическому принципу классифицируют на следующие группы (см. рис.7):
I – преоптическая, в которую входят перивентрикулярное, латеральное и медиальное преоптические ядра
II – передняя, в которую входят супрахиазматическое ядро, супраоптическое и паравентрикулярное ядра
III – средняя, к которой относятся вентромедиальное и дорсомедиальное ядра, ядра воронки
IV – наружная, включающая латеральное гипоталамическое ядро и ядра серого бугра
V – задняя, среди ядер которой различают заднее гипоталамическое, перифорникальное, а также комплекс маммилярных ядер – ядер сосцевидных тел (премаммилярное, супраммамилярное и латеральное маммилярное).
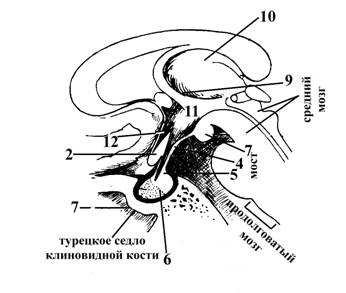
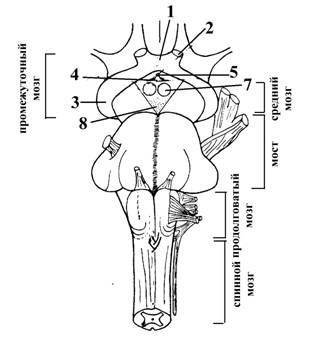
Рис.5. Ствол головного мозга Рис.6. Стволовая часть головного
(вид с вентральной стороны) мозга (сагиттальный разрез).
1 – зрительный перекрест (волокна зрительных нервов, IIп (2) со стороны основания головного мозга большей частью переходят на противоположную сторону, образуя зрительный перекрест, после которого они уже продолжаются в составе зрительных трактов (3)).
3 – зрительный тракт
4 – серый бугор, принимает участие в образовании дна III желудочка головного мозга, переходит в воронку или гипофизарную ножку (5) к которой прикрепляется гипофиз (6).
5 – воронка или ножка гипофиза
6 – гипофиз
7 – сосцевидные тела, находятся между серым бугром (4) и задним продырявленным веществом (8).
9 – подбугорная Монроева борозда, отделяет нижние поверхности таламусов (зрительных бугров) –(10) от гипоталамуса. Область гипоталамуса, находящаяся ниже этой борозды называется субталамической областью (11) и является продолжением ножек мозга (структур среднего мозга); в нее продолжаются и в ней заканчиваются черная субстанция, красное ядро, а также в ней залегает Льюисово тело, которое является звеном экстрапирамидной системы и вегетативным центром.
На рис.6, кроме того, обозначена:
12 – терминальная (пограничная или конечная) пластинка – структура большого (или конечного) мозга, представляющая собой слой серого вещества, связанного своим одним концом со зрительным перекрестом (2), другим – с мозолистым телом большого мозга.
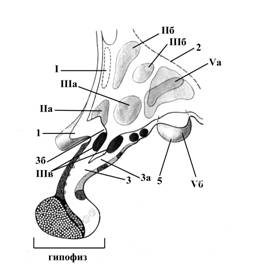
Рис.7. Схема гипоталамуса и проекции его ядер; сагиттальный разрез.
1 – зрительный перекрест
2 – подбугорная Монроева борозда
3 – воронка или гипофизарная ножка
3а – углубление воронки
3б – медиальное возвышение
4 – гипофиз
5 – сосцевидное тело
Ядра гипоталамуса по топографическому принципу классифицируют на следующие группы:
I – преоптическая
II – передняя, в которую входят супрахиазматическое ядро (не показано на рисунке), супраоптическое (IIа) и паравентрикулярное (IIб) ядра
III – средняя, к которой относятся вентромедиальное (IIIа) и дорсомедиальное (IIIб) ядра, ядра воронки (IIIв)
IV – наружная, включающая латеральное гипоталамическое ядро (не показано на рисунке) и ядра серого бугра (IVа)
V – задняя, среди ядер которой различают заднее гипоталамическое, перифорникальное (Vа), а также комплекс маммилярных ядер - ядер сосцевидных тел (премаммилярное, супраммамилярное и латеральное маммилярное) – обозначены цифрой Vа.
Афферентные сигналы в гипоталамус поступают из коры больших полушарий, таламуса, базальных ганглиев большого мозга. Таким образом, функциональное состояние нейронов гипоталамуса, с одной стороны, зависит от той многообразной чувствительной информации, которая достигает таламуса, а с другой – находится под регулирующим контролем большого мозга.
Функциональное значение гипоталамуса
Гипоталамусу отводят роль высшего вегетативного центра в организме в связи с тем, что он, благодаря определенным своим структурам, имеет отношение к регуляции практически всех вегетативных функций. Причем характер регулирующего влияния гипоталамуса на деятельность внутренних органов состоит в том, что он, в отличие от большинства жизненноважных центров (таких как дыхательный, кардиорегуляторный, сосудодвигательный), регулирующих исключительно деятельность какого-то отдельного органа или системы, перестраивает сочетанно деятельность многих физиологических систем в зависимости от текущих потребностей организма. Экспериментально доказано, что латеральная и дорсальная группы ядер гипоталамуса повышают тонус симпатического отдела нервной системы, тогда как средняя – напротив, его снижает. В гипоталамусе находятся центры сна и пробуждения (а следовательно, он имеет отношение к регуляции циркадианных ритмов у человека), голода (латеральные ядра) и насыщения (вентромедиальные ядра), терморегуляции (задние ядра – центр теплопродукции, а передние – центр теплоотдачи), жажды (область, расположенная дорсолатерально от супраоптического ядра), удовольствия и ряд других.
Наряду с непосредственным влиянием гипоталамуса на определенные эфферентные вегетативные нейроны ц. н.с, он участвует в регуляции многих вегетативных функций и косвенно, через посредство эндокринной системы. Причем такая связь гипоталамуса с гуморальными механизмами регуляции физиологических функций достигается, с одной стороны, благодаря его тесным структурно-функциональным взаимоотношениям с гипофизом – дирижером в эндокринной системе, а с другой – путем продукции некоторых гормонов (нейросекретов), обладающих системным действием на организм. В частности, два ядра переднего гипоталамуса (супраоптическое и паравентрикулярное) вырабатывают гормоны (вазпрессин или антидиуретический гормон и окситоцин), которые по аксонам нейронов этих ядер, формирующих гипоталамо-гипофизирный тракт, достигают нейрогипофиза, на его уровне переходят в общий кровоток и оказывают собственные эффекты на организм. Наконец, некоторые ядра медиобазального гипоталамуса (дорсо - и вентромедиальное, аркуатные, вентрикулярные и др.) продуцируют т. н. рилизинг-факторы гипоталамуса, которые поступают в аденогипофиз и оказывают регулирующее влияние на синтез им тропных гормонов. Некоторые из этих тропных гормонов (соматотропный гормон и пролактин) обладают собственными эффектами на организм, а другие (ФСГ, ЛГ, ТТГ, АКТГ) – регулируют секреторную активность ряда периферических эндокринных желез. Благодаря связи гипоталамуса с гипофизом, образующих в совокупности гипоталамо-гипофизрную систему, достигается единство и взаимосвязь нервных и эндокринных механизмов регуляции физиологических функций в организме.
БОЛЬШОЙ (КОНЕЧНЫЙ) МОЗГ
Конечный мозг (telencephalon) состоит из двух полушарий большого мозга, разделенных продольной щелью и соединяющихся между собой в глубине этой щели при помощи мозолистого тела, передней, задней спаек и спайки свода. Полость конечного мозга образуют правый и левый боковые желудочки, каждый из которых находится в соответствующем полушарии.
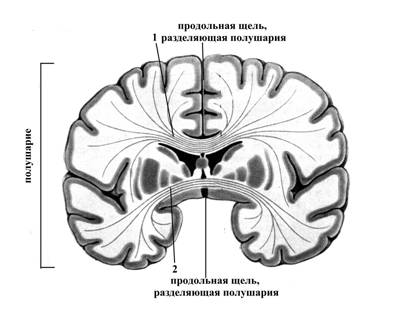
Рис.8. Схематическое изображение фронтального разреза большого мозга
1 – волокна мозолистого тела (большая спайка мозга)
2 – передняя спайка большого мозга
1 и 2 вместе с задней спайкой и спайкой свода (на рисунке не обозначены) соединяют полушария большого мозга друг с другом.
Полушарие большого мозга состоит из:
Ø коры полушария – серого вещества, образующего наружный его покров (плащ большого полушария)
Ø белого вещества, занимающего внутреннее положение в полушарии,
Ø базальных ядер (скоплений серого вещества, расположенных в толще белого вещества).
Поверхность полушарий (их кора) не является гладкой, а имеет довольно сложный рельеф, связанный с наличием более или менее глубоких борозд и расположенных между ними валикообразных возвышений – извилин большого мозга. Причем 2/3 коры большого мозга находится в бороздах и 1/3 – в извилинах. Наличие в коре борозд и извилин увеличивает суммарную площадь ее поверхности.
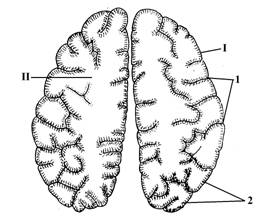
Рис.9. Полушария большого мозга на поперечном разрезе
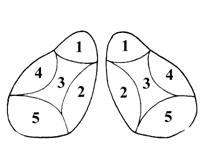
I – кора больших полушарий (плащ)
1 – борозды (углубления в коре)
2 – извилины (участки коры между бороздами).
II – белое вещество
Каждое полушарие имеет три поверхности:
Ø верхнелатеральную (наиболее выпуклую)
Ø медиальную (плоскую, обращенную к соседнему полушарию)
Ø нижнюю (имеет сложный рельеф, соответствующий внутреннему основанию черепа).
Поверхности полушарий отделены друг от друга краями: верхним, нижнелатеральным и нижнемедиальным. Наиболее выступающие участки полушарий получили название полюсов. В частности, в каждом полушарии выделяют три полюса:
Ø лобный полюс
Ø затылочный полюс
Ø височный полюс.
Несмотря на то, что глубина, протяженность, форма и направление борозд и извилин очень изменчивы, локализация и форма основных глубоких борозд и извилин у различных людей более или менее постоянны. Глубокие борозды коры больших полушарий разделяют их на доли. Так, в больших полушариях выделяют четыре доли:
Ø лобная (находится в переднем отделе полушарий, заканчивается спереди лобным полюсом, отделена от расположенной ниже ее височной доли латеральной (сильвиевой) бороздой, а от находящейся позади ее теменной доли – центральной бороздой)
Ø теменная (находится позади лобной доли, от которой отделена центральной бороздой; нижней ее границей (границей с височной долей) является сильвиева борозда, а задней границей (границей с затылочной долей) – теменно-затылочная борозда)
Ø затылочная (имеет небольшие размеры, располагается позади теменной доли и заканчивается затылочным полюсом)
Ø височная (занимает нижнебоковые отделы в полушариях, отделена от лобной и теменной долей сильвиевой бороздой, передняя часть этой доли выпуклая и образует височный полюс)
Ø островковая (находится в глубине сильвиевой борозды, снаружи прикрыта участками лобной, теменной и височной долей).

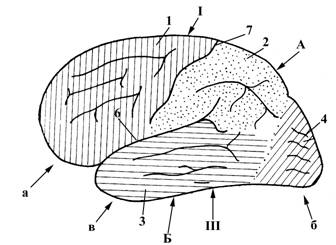
Рис.10. Полушарие большого мозга, Рис.11. Полушарие большого мозга,
вид с латеральной стороны вид с латеральной стороны (часть лобной, теменной
 и височной долей удалена)
и височной долей удалена)
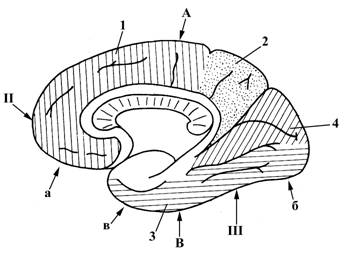
Рис.12. Полушарие большого мозга, вид с медиальной стороны
В больших полушариях выделяют три поверхности:
ü верхнелатеральная (I), выпуклая
ü медиальная (II), плоская
ü нижняя (III).
Эти три поверхности отделены друг от друга краями:
ü верхним (А), отделяющим верхнелатеральную поверхность (I) от нижней (III);
ü нижнелатеральным (Б), отделяющим верхнелатеральную поверхность (I) от нижней (III);
ü нижнемедиальным (В), отделяющим медиальную поверхность(III) от нижней (II).
Наиболее выступающие поверхности полушарий носят название полюсов. Выделяют 3 полюса:
а – лобный
б – затылочный
в – височный.
Поверхность полушарий не является гладкой, она разделяется большим количеством борозд (углублений) на извилины; последние объединяются в группы, разделенные более глубокими бороздами, и называемые долями. В каждом полушарии выделяют следующие доли:
1 – лобная
2 – теменная
3 – височная
4 – затылочная
5 – островковая доля (находится в глубине латеральной борозды (6), прикрыта лобной, височной и теменной долями).
Лобная доля (1) отделена от теменной (2) центральной (Ролландовой) бороздой (7); лобная (1) и теменная (2) доли отделены от височной (3) латеральной бороздой (6).
Цитоархитектоника (клеточная структура) коры больших полушарий
Кора больших полушарий является филогенетически наиболее молодым образованием мозга. В структурном отношении она представляет собой слой серого вещества, образованного нервными клетками и покрывающего большие полушария. Ее толщина в различных участках больших полушарий варьирует от 1,5 до 5мм; общая поверхность коры обоих полушарий у взрослого человека составляет от 1450 до 1700см2 (площадь поверхности коры одного полушария в среднем – 220тыс. мм2), а общее количество нейронов составляющих кору – 12-18 млрд.
В соответствии с особенностями происхождения различных участков коры больших полушарий в филогенезе, различают следующие три ее типа:
Ø древняя кора (архикортекс), включающая структуры проводникового отдела обонятельного анализатора (обонятельные луковицы (образованные совокупностью нейронов, получающих информацию от обонятельного эпителия носовой полости по волокнам обонятельного нерва), обонятельные тракты (расположенные на нижней поверхности лобной доли), обонятельные бугорки и обонятельные треугольники);
Ø старая кора (палеокортекс), включает некоторые области медиальной поверхности полушарий (поясную извилин и извилину гиппокампа)
Ø новая кора (неокортекс), объединяющая все остальные области коры больших полушарий головного мозга.
Несмотря на значительную функциональную специализацию различных областей коры, их нейронная структура имеет много общего. В частности, в коре больших полушарий млекопитающих животных, в том числе человека, выделяют 6 слоев последовательно уложенных (друг над другом) и определенным образом взаимосвязанных между собой нервных элементов:
I слой – молекулярный (занимает самое поверхностное положение в коре), беден нервными клетками, образован в основном отростками нейронов нижележащих слоев (в частности, в этом слое многократно разветвляются дендриты нейронов V слоя – дендриты крупных пирамидных клеток);
II слой – наружный зернистый, образован мелкими густо расположенными клетками-зернами (диаметр около 4-8мкм), имеющими округлую, треугольную или многоугольную форму;
III слой – наружный пирамидный, более толстый, чем первые два слоя, образован пирамидными и звездчатыми нейронами различных размеров;
IV слой – внутренний зернистый, подобно II слою образован скоплением мелких клеток-зерен;
V слой – внутренний пирамидный, образован крупными пирамидными нейронами Беца (диаметр до 125 мкм), характеризуется наибольшей толщиной и размерами клеток в области моторных зон коры; дендриты пирамидных нейронов этого слоя достигают поверхности коры (молекулярного слоя), где образуют густые разветвления, а аксоны пирамидных нейронов Беца формируют единственный эфферентный выход из коры больших полушарий и направляются к базальным ганглиям конечного мозга, ядрам ствола головного мозга и нейронам спинного мозга.
VI слой – мультиформный, образован веретенообразными и треугольной формы нейронами.
По функциональному принципу все нейроны коры могут быть разделены на три группы:
Ø сенсорные, воспринимают чувствительную информацию, поступающую в кору по афферентным ее входам (преимущественно аксонам нейронов ядер таламуса и базальных ганглиев); таковыми в коре преимущественно являются клетки III (наружный пирамидный) и IV (внутренний зернистый) слоев в сенсорных областях коры;
Ø эффекторные, аксоны которых образуют эфферентный выход из коры больших полушарий и направляются к нейронам базальных ганглиев, мозгового ствола или спинного мозга; таковыми являются крупные пирамидные нейроны V слоя коры (внутреннего пирамидного), преимущественно заложенные в моторных областях коры;
Ø контактные (промежуточные) нейроны, обеспечивают связь между различными корковыми нейронами; таковыми являются преимущественно веретенообразные нейроны II (наружный зернистый) и VI (мультиформный) слоев.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
