

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Нарушение сбалансированности систем, контролирующих нормальное развитие отдаленных гибридов, может привести к нарушениям в мейозе при расхождении хромосом в анафазе и образованию гамет с разным уровнем плоидности. Поэтому мы вправе предположить, что триплоиды и тетраплоиды могут формировать женские гаметы разных типов (рис.2).
Таблица 2 - Результаты использования метода эмбриокультуры при получении сеянцев F2 отдаленных гибридов
Показатель | Происхождение зародышей | |||
триплоид х тетраплоид | тетраплоид х гексаплоид | тетраплоид х (диплоид + гексаплоид) | тетраплоид х тетраплоид | |
Средняя оценка развития зародышей, балл | 5,0 | 4,2 | 4,6 | 4,5 |
Посеяно зародышей, шт. | 2 | 35 | 8 | 21 |
Развились в проростки, шт. % | 2 100 | 30 85,7 | 8 100 | 20 95,2 |
Высажено сеянцев в грунт, шт. % | 1 50 | 27 77,1 | 7 87,5 | 17 81,0 |
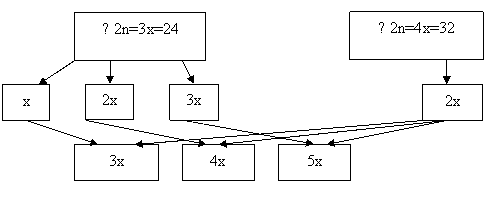
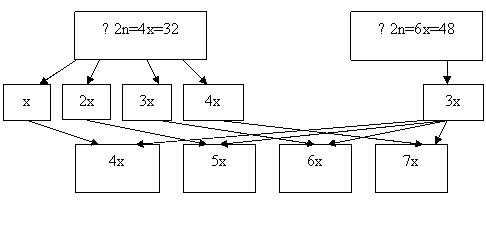
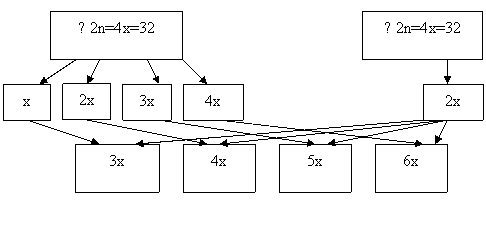
Первая группа гибридов – х=8, 2х=16, 3х=24, вторая – х=8, 2х=16, 3х=24, 4х=32. При гибридизации триплоидов (F1) с терном в результате свободного комбинирования возможных типов гамет можно ожидать появление следующих классов потомства (F2): 3х=24, 4х=32, 5х=40. В комбинации скрещивания 4х (F1) х 6х : 4х=32, 5х=40, 6х=48, 7х=56. При опылении 4х (F1) х 4х, могут сформироваться гибридные организмы, имеющие кариотип: 3х=24, 4х=32, 5х=40, 6х=48. Вполне возможно появление анеуплоидных форм (моносомиков, нуллисомиков, трисомиков).
Исследования показали, что сеянцы F2 от скрещивания 4х (96-68) х 6х, 4х (96-68) х (2х+6х) и 4х х 4х характеризуются низкой степенью подмерзания (0-1 балл). Вместе с тем, в гибридной семье 4х х 4х одиночные растения имели 4-5 баллов подмерзания, несмотря на то (по литературным данным), что терн обладает высокой зимостойкостью и передает этот признак гибридам. Очевидно, при отдаленной гибридизации формируется широкий спектр форм по наследованию определенного признака. Исследованиями установлено, что у некоторых гибридных растений (F2) отмечали ограниченную силу роста. Причем, различия в силе роста между отдельными сеянцами в пределах гибридной семьи наблюдали уже по мере их роста в вазонах с почвой. По-нашему мнению, это связано с тем, что одним из компонентов скрещивания при получении F1 и F2 являлись слаборослые формы – сорт вишнесливы Опата и терн, которые могли передать сеянцам гены, контролирующие этот признак.
триплоид (F1) тетраплоид (терн)
| а б в | |
гетраплоид (F1) гексаплоид (слива домашняя) тетраплоид (F1) тетраплоид (терн)
| а б в | |
Рис. 3. Формообразование в комбинациях скрещивания с участием гибридов сливы (F1): а – исходные формы; б – гаметы; в – гибрид (F2). Х-основное число хромосом рода Prunus Mill равно 8.
| а б в |
Так, в гибридных семьях 4х (96-68) х 6х и 4х (96-68) х (2х+6х) выделены формы с естественным сдержанным ростом, высота которых в пятилетнем возрасте не превышала 1 м.
4.2. Влияние физиологически активных веществ на развитие зародышей яблони в культуре in vitro. Исследованиями установлено, что введение в питательную среду экзогенных результатов роста оказывало положительное влияние на формирование проростков. В ряде вариантов опыта наблюдалось развитие из каждого зародыша нескольких побегов (табл.3).
Таблица 3 - Влияние экзогенных регуляторов роста на развитие
изолированных зародышей (среда Мурасиге-Скуга)
Происхождение зародышей (сорт) | Концентрация фитогормонов, мг/л | Число зародышей в опыте, шт. | Развились в проростки, шт. | Всего сформировалось побегов, шт. | Среднее число побегов из зародыша, шт. |
Ренет Черненко | 0 (контроль) | 20 | 2 | 2 | 1,0 |
БАП 1,0 | 16 | 10 | 26 | 2,6 | |
БАП 2,0 | 22 | 12 | 76 | 6,3 | |
БАП 4,0 | 37 | 11 | 79 | 7,2 | |
БАП 1,6 + ГК 0,5 + ИМК 0,1 | 20 | 7 | 7 | 1,0 | |
НСР05 | 3,0 | ||||
Кандиль Горшкова | 0 (контроль) | 40 | 5 | 5 | 1,0 |
БАП 1,0 | 44 | 24 | 48 | 2,0 | |
БАП 2,0 | 41 | 23 | 64 | 2,8 | |
БАП 3,0 | 41 | 13 | 89 | 6,8 | |
БАП 4,0 | 41 | 7 | 20 | 2,9 | |
БАП 1,6 + ГК 0,5 + ИМК 0,1 | 40 | 28 | 28 | 1,0 | |
НСР05 | 1,28 | ||||
Пепин шафранный | 0 (контроль) | 40 | 3 | 3 | 1,0 |
БАП 1,0 | 36 | 8 | 17 | 2,1 | |
БАП 2,0 | 30 | 6 | 6 | 1,0 | |
БАП 3,0 | 36 | 14 | 38 | 2,7 | |
БАП 1,6 + ГК 0,5 + ИМК 0,1 | 15 | 5 | 5 | 1,0 | |
НСР05 | 0,93 | ||||
Антоновка обыкновенная | 0 (контроль) | 35 | 4 | 4 | 1,0 |
БАП 1,0 | 36 | 8 | 26 | 3,3 | |
БАП 2,0 | 30 | 5 | 5 | 1,0 | |
БАП 3,0 | 34 | 6 | 30 | 5,0 | |
БАП 1,6 + ГК 0,5 + ИМК 0,1 | 30 | 8 | 8 | 1,0 | |
НСР05 | 1,26 | ||||
Грушовка московская | 0 (контроль) | 30 | 0 | 0 | 0 |
БАП 1,0 | 28 | 8 | 21 | 2,6 | |
БАП 2,0 | 33 | 7 | 16 | 2,3 | |
БАП 3,0 | 31 | 9 | 28 | 3,1 | |
БАП 1,6 + ГК 0,5 + ИМК 0,1 | 22 | 6 | 6 | 1,0 | |
НСР05 | 1,21 |
Способность зародышей к пролиферации дополнительных побегов отмечалась при добавлении в питательную среду 6-БАП, что связано с её физиологическим эффектом снимать апикальное доминирование верхушечной почки в растении. Воздействуя на верхушечную почку зародыша, 6-БАП вызывает дифференциацию пазушных почек, их пробуждение и активацию роста в побеги. Зародыши по-разному реагировали на присутствие в среде цитокинина. При этом эффект действия зависел от его концентрации, комбинации с другими регуляторами роста генотипических особенностей. Менее интенсивно протекал процесс активации прорастания пазушных почек и развитие побегов у зародышей культивируемых на других испытанных вариантов сред и в основном при сочетании 6-БАП с ИУК или ИМК. Эти варианты были на уровне контроля.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
