

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
4.2. ОБЩИЕ ЗАКОНОМЕРНОСТИ ВОЗДЕЙСТВИЯ
ЭКОЛОГИЧЕСКИХ ФАКТОРОВ НА ЖИВЫЕ ОРГАНИЗМЫ
Несмотря на большое разнообразие экологических факторов, в характере их воздействия на организмы и в ответных реакциях живых существ можно выявить ряд общих закономерностей.
1. Закон оптимума. Каждый фактор имеет лишь определенные пределы положительного влияния на организмы. Результат действия переменного фактора зависит прежде всего от силы его проявления. Как недостаточное, так и избыточное действие фактора отрицательно сказывается на жизнедеятельности особей. Благоприятная сила воздействия называется зоной оптимума экологического фактора или просто оптимумом для организмов данного вида. Чем сильнее отклонение от оптимума, тем больше выражено угнетающее действие данного фактора на организмы (зона пессимума). Максимально или минимально переносимые значения фактора – это критические точки, за пределами которых существование уже невозможно, наступает смерть. Пределы выносливости между критическими точками называют экологической валентностью живых существ по отношению к конкретному фактору среды.
Представители разных видов сильно отличаются друг от друга как по положению оптимума, так и по экологической валентности. Одна и та же сила проявления фактора может быть оптимальной для одного вида, пессимальной для другого и выходить за пределы выносливости для третьего.
Широкую экологическую валентность вида по отношению к абиотическим факторам среды обозначают добавлением к названию фактора приставки «эври».
Эвритермные виды – выносящие значительные колебания температуры, эврибатные – широкий диапазон давления, эвригалинные – разную степень засоления среды.
Неспособность переносить значительные колебания фактора или узкая экологическая валентность характеризуются приставкой «стено»: стенотермные, стенобатные, стеногалинные виды и т. д. В более широком смысле слова виды, для существования которых необходимы строго определенные экологические условия, называются стенобатными, а те, которые способны приспосабливаться к разной экологической обстановке – эврибатными.
2. Неопределенность действия фактора на разные функции. Каждый фактор неодинаково влияет на разные функции организма. Оптимум для одних процессов может являться пессимумом для других. Для многих рыб температура воды, оптимальная для созревания половых продуктов, неблагоприятна для икрометания, которое происходит при другом температурном интервале.
Жизненный цикл, в котором в определенные периоды организм осуществляет преимущественно те или иные функции (питание, рост, размножение, расселение и т. п.), всегда согласован с сезонными изменениями комплекса факторов среды. Подвижные организмы могут также менять места обитания для успешного осуществления всех своих жизненных функций.
3. Изменчивость, вариабельность и разнообразие ответных реакций на действия факторов среды у отдельных особей вида. Степень выносливости, критические точки, оптимальные и пессимальные зоны отдельных индивидуумов не совпадают. Эта изменчивость определяется как наследственными качествами особей, так и половыми, возрастными и физиологическими различиями. Например, у бабочки ельничной огневки (одного из вредителей муки и зерновых продуктов) критическая минимальная температура для гусениц – 7 °С, для взрослых форм – 10 °С, а для яиц – 27 °С. Мороз в 10 °С губит гусениц, но не опасен для яиц этого вредителя. Следовательно, экологическая валентность вида всегда шире экологической валентности отдельной особи.
4. К каждому из факторов среды виды приспосабливаются относительно независимым путем. Степень выносливости к какому-нибудь фактору не означает соответствующей экологической валентности вида по отношению к остальным факторам. Например, виды, переносящие широкие изменения температуры, совсем не обязательно должны также быть приспособлены к широким колебания влажности или солевого режима. Эвритермные виды могут быть стеногалинными, стенобатными или наоборот. Экологические валентности вида по отношению к разным факторам могут быть очень разнообразными. Это создает чрезвычайное многообразие адаптаций в природе. Набор экологических валентностей по отношению к разным факторам среды составляет экологический спектр вида.
5. Несовпадение экологических спектров отдельных видов. Каждый вид специфичен по своим экологическим возможностям. Даже у близких по способам адаптации к среде видов существуют различия в отношении к каким-либо отдельным факторам.
Правило экологической индивидуальности видов сформулировал русский ботаник применительно к растениям, а затем это было подтверждено и зоологическими исследованиями.
6. Взаимодействие факторов. Оптимальная зона и пределы выносливости организмов по отношению к какому-либо фактору среды могут смещаться в зависимости от того, с какой силой и в каком сочетании действуют одновременно факторы. Эта закономерность получила название взаимодействия факторов. Например, жару легче переносить в сухом, а не во влажном воздухе. Таким образом, один и тот же фактор в сочетании с другими оказывает неодинаковое экологическое воздействие. Наоборот, один и тот же экологический результат может быть получен разными путями. Например, увядание растений можно приостановить как увеличением количества влаги в почве, так и снижением температуры воздуха, уменьшающего испарение. Создается эффект частичного взаимозамещения факторов.
Вместе с тем взаимная компенсация действия факторов среды имеет определенные пределы, и полностью заменить один из них другим нельзя. Полное отсутствие воды или хотя бы одного из элементов минерального питания делает жизнь растения невозможной, несмотря на самые благоприятные сочетания других условий.
Если хотя бы один из экологических факторов приближается или выходит за пределы критических величин, то, несмотря на оптимальное сочетание остальных условий, особям грозит гибель. Также сильно уклоняющиеся от оптимума факторы приобретают первостепенное значение в жизни вида или отдельных его представителей в каждый конкретный отрезок времени.
Лимитирующий фактор (условие) – это любой фактор (условие), приближающийся к пределу толерантности или превышающий его.
Толерантность экологическая есть способность организма переносить отклонения экологических факторов от оптимальных для себя; область устойчивого существования вида или реализации какой-либо его функции по отношению к конкретному фактору среды или их сочетанию.
Устойчивость вида – способность вида сохранять функционирование в пределах естественного колебания его параметров.
Гомеостаз – состояние внутреннего динамического равновесия природной системы (организма, вида), поддерживаемого регулярным возобновлением его основных структур, вещественно-энергетического состава и постоянной функциональной саморегуляции ее компонентов.
Саморегуляция – свойство системы в процессе функционирования сохранять на определенном уровне типичное состояние, режимы, характеристику связей между ее компонентами.
Компоненты природной системы – основные составные её части, тесно связанные между собой; при изменении одного из компонентов меняются другие и возникают «цепные реакции».
Ограничивающие факторы среды определяют географический ареал вида. Природа этих факторов может быть различной. Так, продвижение вида на север может лимитироваться недостатком тепла, в аридные районы – недостатком влаги или слишком высокими температурами. фактором, ограничивающим распространение, могут служить биотические отношения, например занятость территории более сильным конкурентом или недостаток опылителей для растений.
Закон Либиха (закон минимума): «веществом, присутствующим в минимуме, управляется урожай» – и закон Шелфорда (закон толерантности): «любой организм имеет верхние и нижние границы устойчивости (толерантности) к любому экологическому фактору» – являются фундаментальными основополагающими законами экологии.
Контрольные вопросы
1. Что понимается под средой обитания?
2. В чём разница между биотическими и абиотическими факторами среды?
3. Перечислите физические и химические факторы неживой природы.
4. Что относится к климатическим факторам?
5. Что изучает наука геоморфология?
6. Что является отличительной особенностью антропогенных факторов? Перечислите основные.
7. Перечислите основные законы воздействия экологических факторов на живые организмы.
8. Что такое экологическая толерантность?
9. Сформулируйте закон Либиха (закон минимума).
10. Что такое гомеостаз?
Глава V. ЭКОСИСТЕМЫ
5.1. ПОТОК ЭНЕРГИИ И КРУГОВОРОТ
ПИТАТЕЛЬНЫХ ВЕЩЕСТВ
При изучении основных понятий экологии мы выяснили, что термин «экосистемы» означает не что иное, как «сообщество живых организмов и среды обитания, составляющее единое целое на основе пищевых связей и способов получения энергии». Термин ввёл А. Тэнсли в 1935 г. При изучении данной главы мы познакомимся с «работой экосистемы», т. е. в общем виде с тем, как живые организмы получают энергию, которая позволяет им жить и существовать в том сообществе, которое составляет единое целое – экосистему.
Организмы в экосистеме связаны общностью энергии и питательных веществ, и необходимо четко разграничить эти два понятия.
Энергия определяется как способность совершать работу. Живые организмы нуждаются в энергии для поддержания жизни точно так же, как машины для того, чтобы работать. Всю экосистему можно уподобить единому механизму, потребляющему энергию и питательные вещества для совершения работы. Питательные вещества первоначально происходят из абиотического компонента системы, в который, в конце концов, и возвращаются либо в качестве отходов жизнедеятельности, либо после гибели и разрушения организмов. Таким образом, в экосистеме происходит постоянный круговорот питательных веществ, в котором участвуют живые и неживые компоненты. Такие круговороты называются биогеохимическими циклами.
Движущей силой этих круговоротов служит, в конечном счете, энергия Солнца. Фотосинтезирующие организмы непосредственно используют энергию солнечного света и затем передают ее другим представителям биотического компонента. В итоге создается поток энергии и питательных веществ через экосистему, как показано на рис. 3.

Рис. 3. Поток энергии и круговороты биогенных элементов в экосистеме
Необходимо отметить, что климатические факторы абиотического компонента, такие как температура, движение атмосферы, испарение и осадки, тоже регулируются поступлением солнечной энергии.
Для того чтобы понять, почему имеет место линейный поток энергии через экосистему, а не ее круговорот и повторное использование (как в случае питательных веществ), необходимо коротко рассмотреть некоторые термодинамические положения.
Энергия может существовать в виде различных взаимопревращаемых форм, таких как механическая, тепловая и электрическая энергия. Переход одной формы в другую, называемый преобразованием энергии, подчиняется законам термодинамики. Первый закон термодинамики, закон сохранения энергии, гласит, что энергия может превращаться из одной формы в другую, но не может быть создана или уничтожена. Второй закон утверждает, что при совершении работы энергия не может быть использована на все 100 % и часть ее неизбежно превращается в тепло. Тепло есть результат случайного движения молекул, тогда как работа всегда означает неслучайное (т. е. упорядоченное) использование энергии. Понятие «работы» приложимо к любому процессу, протекающему в живой системе с потреблением энергии, начиная от процессов на клеточном уровне, таких как поддержание электрических градиентов на мембране и синтез белков, и кончая процессами на уровне целого организма (рост, развитие, размножение).
Таким образом, живые организмы – это преобразователи энергии, и каждый раз, когда происходит превращение энергии, часть ее теряется в виде тепла. В конце концов, вся энергия, поступающая в биотический компонент экосистемы, рассеивается в виде тепла. Можно было бы подумать, что поскольку и тепло способно совершать работу (например, в паровозе), то нет причин, которые мешали бы круговороту тепла. Однако процесс, производящий тепло, требует больше энергии, чем может быть возвращено путем вторичного использования этого тепла, поэтому в целом происходит все же потеря полезной энергии в системе. Фактически живые организмы не используют тепло как источник энергии для совершения работы – они используют свет и химическую энергию.
Изучение потока энергии через экосистемы называется энергетикой экосистем, и так как это доминирующая тема в их изучении, нам важно познакомиться с единицами, используемыми для измерения энергии.
5.2. ПИЩЕВЫЕ ЦЕПИ И ТРОФИЧЕСКИЕ УРОВНИ
Внутри экосистемы содержащие энергию органические вещества создаются автотрофными организмами и служат пищей (источником вещества и энергии) для гетеротрофов. Типичный пример: животное поедает растения. Это животное в свою очередь может быть съедено другим животным, и таким путем может происходить перенос энергии через ряд организмов – каждый последующий питается предыдущим, поставляющим ему сырье и энергию. Такая последовательность называется пищевой цепью, а каждое ее звено – трофическим уровнем (греч. «Trophos» – питание). Первый трофический уровень занимают автотрофы, или, так называемые, первичные продуценты. Организмы второго трофического уровня называются первичными консументами, третьего – вторичными консументами и т. д. Обычно бывает четыре или пять трофических уровней и редко больше шести.
Первичные продуценты. Первичными продуцентами являются автотрофные организмы, в основном зеленые растения, некоторые прокариоты, а именно сине-зеленые водоросли и немногочисленные виды бактерий, которые тоже фотосинтезируют, но их вклад относительно не велик. Фотосинтетики превращают солнечную энергию (энергию света) в химическую энергию, заключенную в органических молекулах, из которых построены их ткани. Небольшой вклад в продукцию органического вещества вносят и хемосинтезирующие бактерии, извлекающие энергию из неорганических соединений.
В водных экосистемах главными продуцентами являются водоросли – часто мелкие одноклеточные организмы, составляющие фитопланктон поверхностных слоев океана и озер. На суше большую часть первичной продукции поставляют более организованные формы, относящиеся к голосеменным и покрытосеменным. Они формируют леса и луга.
Первичные консументы. Первичные консументы питаются первичными продуцентами, то есть травоядные животные. На суше типичными травоядными являются многие насекомые, рептилии, птицы и млекопитающие. Наиболее важные группы травоядных млекопитающих – это грызуны и копытные. К последним относятся пастбищные животные (такие как лошади, овцы, крупный рогатый скот), приспособленные к бегу на кончиках пальцев.
В водных экосистемах (пресноводных и морских) травоядные формы представлены обычно моллюсками и мелкими ракообразными. Большинство этих организмов – ветвистоусые и веслоногие рачки, личинки крабов, усоногие раки и двустворчатые моллюски (например, мидии и устрицы) – питаются, отфильтровывая мельчайших первичных продуцентов из воды.
Консументы второго и третьего порядка. Вторичные консументы питаются травоядными; таким образом, это уже плотоядные животные, так же как и третичные консументы, поедающие консументов второго порядка. Консументы второго и третьего порядка могут быть хищниками и охотиться, схватывать и убивать свою жертву, могут питаться падалью или быть паразитами. В последнем случае они по величине меньше своих хозяев.
Редуценты и детритофаги (детритные пищевые цепи). Существуют два главных типа пищевых цепей – пастбищные и детритные. Выше мы рассмотрели примеры пастбищных цепей, в которых первый трофический уровень занимают зеленые растения, второй – пастбищные животные (термин «пастбищные» используется в широком смысле и включает все организмы, питающиеся растениями) и третий – хищники. Тела погибших растений и животных еще содержат энергию и «строительный материал» так же, как и прижизненные выделения, например, моча и фекалии. Эти органические материалы разлагаются микроорганизмами, а именно грибами и бактериями, живущими как сапрофиты на органических остатках. Такие организмы называются редуцентами. Они выделяют пищеварительные ферменты на мертвые тела или отходы жизнедеятельности и поглощают продукты их переваривания. Скорость разложения может быть различной.
Кусочки частично разложившегося материала называют детритом, и многие мелкие животные (детритофаги) питаются им, ускоряя процесс разложения. Поскольку в этом процессе участвуют как истинные редуценты (грибы и бактерии), так и детритофаги (животные), и тех и других иногда называют редуцентами, хотя в действительности этот термин относится только с сапрофитным организмам.
В схемах пищевых цепей каждый организм бывает представлен как питающийся другими организмами какого-то одного типа. Однако реальные пищевые связи в экосистеме намного сложнее, так как животное может питаться организмами разных типов из одной и той же пищевой цепи или даже из разных пищевых цепей. Это в особенности относится к хищникам верхних трофических уровней. Некоторые животные питаются как другими животными, так и растениями, их называют всеядными (таков, в частности, человек). В действительности пищевые цепи переплетаются таким образом, что образуется пищевая (трофическая) сеть.
Для изучения взаимоотношений между организмами в экосистеме и для графического представления этих взаимоотношений удобнее использовать не схемы пищевых цепей, а экологические пирамиды. При этом сначала подсчитывают число различных организмов на данной территории, сгруппировав их по трофическим уровням. После таких подсчетов становится очевидным, что численность животных прогрессивно уменьшается при переходе от второго трофического уровня к последующим. Численность растений первого трофического уровня тоже нередко превосходит численность животных, составляющих второй уровень. Это можно отобразить в виде пирамиды численности.
Для удобства количество организмов на данном трофическом уровне может быть представлено в виде прямоугольника, длина которого пропорциональна числу организмов, обитающих на данной площади (или в данном объеме, если это водная экосистема).
Хищники, расположенные на высшем трофическом уровне, называются конечными хищниками. Существует ряд неудобств, связанных с использованием этих пирамид.
Избежать указанные неудобства можно путем построения пирамид биомассы, в которых учитывается суммарная масса организмов (биомасса) каждого трофического уровня. Определение биомассы включает не только учет численности, но и взвешивание отдельных особей, так что это более трудоемкий процесс, требующий больше времени и специального оборудования.
Наиболее фундаментальным и в определенном смысле идеальным способом отображения связи между организмами на разных трофических уровнях служит пирамида энергии, обладающая рядом преимуществ:
1. Она отражает скорость увеличения биомассы в отличие от пирамид численности и биомассы, описывающих только текущее состояние организмов в отдельный момент времени. Каждая ступенька пирамиды энергии отражает количество энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за определенный период.
2. При одинаковой биомассе два вида совсем не обязательно содержат одинаковое количество энергии. Поэтому сравнение, основанное на биомассе, может ввести в заблуждение.
3. Пирамиды энергии позволяют сравнивать не только различные экосистемы, но и относительную значимость популяций внутри одной экосистемы, не получая при этом перевернутых пирамид.
4. К основанию пирамиды энергии можно добавить еще один прямоугольник, отображающий поступление солнечной энергии.
Хотя пирамиды энергии – наиболее полезный метод из трех перечисленных типов экологических пирамид, получать данные для их построения труднее всего, т. к. при этом требуется даже больше измерений, чем при построении пирамид биомассы. В частности, необходима дополнительная информация об энергетической стоимости данных масс организмов, а это требует сжигания репрезентативных выборок. На практике же пирамиды энергии иногда могут быть с достаточной достоверностью получены из пирамид биомассы путем пересчета, основанного на проведенных ранее экспериментах.
Продуктивность. Скорость накопления энергии первичными продуцентами в форме органического вещества, которое может быть использовано в пищу, называется первичной продуктивностью. Это важный параметр, т. к. им определяется общий поток энергии через биотический компонент экосистемы, а значит и количество (биомасса) живых организмов, которые могут существовать в экосистеме. Количество солнечного света, падающего на растения, различно. Оно завит от географической широты и от степени развития растительного покрова. Примерно вит от географической широты и от степени развития растительного покрова. компонент экосистемы, а значит, и количество ()трофи 95–99 % этого количества сразу отражается, поглощается с переходом в тепло или расходуется на испарение воды, и лишь 1–5 % поглощается хлорофиллом и используется для синтеза органических молекул. Скорость, с которой растения накапливают химическую энергию, называется валовой первичной продуктивностью (ВПП). Это энергия, которую могут использовать организмы следующих трофических уровней.
Животные, как и растения, теряют часть энергии при дыхании. Энергия, оставшаяся после потерь, связанных с процессами дыхания, пищеварения, экскреции, идет на рост, поддержание жизнедеятельности и размножение.
Количество органического вещества, накопленного гетеротрофными организмами, называется вторичной продукцией (на любом трофическом уровне). Приведенное ниже словесное уравнение суммирует расход потребленной животным энергии:
Потребленная пища = рост + дыхание + фекалии + экскреты
Некоторые составляющие этого уравнения не трудно определить на домашних или диких животных в лабораторных условиях. Показателем роста за какое-то время служит увеличение биомассы или же энергии, заключенной в теле. Фекалии и экскреты можно собрать, взвесить и вычесть их массу из общей массы потребленной пищи, чтобы найти массу усвоенной пищи, использованной для роста и других процессов.
Одна из причин, которые побуждают изучать поток энергии, проходящей через экосистему, – это возможность применения полученных знаний для удовлетворения потребности человека в пище и энергии. Научный анализ позволяет оценивать традиционные методы агротехники и вносить усовершенствования с целью повышения их эффективности.
Рациональное использование экосистемы. «Получение урожая» означает изъятие из экосистемы тех организмов или их частей, которые используются в пищу (или для других целей). При этом желательно, чтобы экосистема производила пригодную для пищи продукцию наиболее эффективно. Это может быть достигнуто путем повышения урожайности культуры, уменьшения заболеваемости и помех со стороны других организмов или использования культуры, более приспособленной к условиям данной экосистемы.
Уменьшение вреда от насекомых и птиц, а также от болезней достигается выведением генетически устойчивых сортов и применением химических средств, уничтожающих возбудителей болезней и вредителей. Однако такие средства нужно использовать осторожно. Применение стойких препаратов, неспособных быстро разлагаться в экосистеме, может иметь катастрофические последствия на других трофических уровнях. Частое применения эффективного пестицида может вызывать вспышки численности устойчивых к нему форм вредителей, с которыми трудно бороться. Такая ситуация возникла в Африке при попытках контролировать численность малярийного комара с помощью ДДТ (дихлордифенилтрихлорэтан).
Большую роль при разведении животных, достигающих половой зрелости через несколько лет, играет концепция максимального постоянного уровня добычи. Это наибольшая скорость изъятия особей из популяции без ущерба для ее дальнейшей продуктивности. Для определения этой величины необходимо иметь ясное представление о структуре и динамике популяции, особенно о рождаемости: ведь чем выше рождаемость, тем больше особей можно изымать из популяции. Знание структуры популяции позволяет установить, какие возрастные группы играют наибольшую роль в воспроизводстве стада, и соответственно отобрать для убоя или отлова животных других возрастных групп. В Англии, например, официально установлен минимальный размер ячеек для сетей, которыми можно пользоваться в промысловом рыболовстве. Таким образом, вылавливается более крупная и старая рыба, которая уже дала потомство, а не достигшие половой зрелости особи в сеть, как правило, не попадают. Вылов молоди привел бы к уменьшению воспроизводства рыбного стада в последующие годы и уменьшил бы размеры популяций в будущем.
Эффекты концентрации веществ в пищевых цепях. После второй мировой войны стало быстро возрастать число различных синтетических веществ, выбрасываемых в окружающую среду. Это прежде всего гербициды и пестициды, предназначенные для уничтожения организмов (особенно сорняков), наносящих вред урожаям, домашнему скоту и самому человеку. Среди первых применявшихся с успехом пестицидов была группа хлорированных углеводородов, в т. ч. ДДТ, диэлдрин и алдрин. Эти вещества ядовиты для многих животных и человека, но особенно вредоносны для птиц, рыб и беспозвоночных (см. рис. 4).
Птица | 75 | Хищник 2 |
Крупная рыба | 50 | Хищник 1 |
Мелкая рыба | 10 | Травоядные животные |
Водные растения | 0,04 | Продуцент |
Рис. 4. Количество ДДТ, заключенное в биомассе организмов, находящихся на разных трофических уровнях пищевой цепи
Цифрами выражено количество весовых единиц ДДТ, приходящихся на 1 млн. весовых единиц биомассы.
В середине 60-х годов неожиданным для многих ученых явилось сообщение о том, что ДДТ обнаружен в печени пингвинов в Антарктиде, весьма удаленной от районов возможного применения ДДТ.
От отравления пестицидами очень сильно пострадали некоторые хищники верхних трофических уровней, большего всего птицы. Например, из-за отравления ДДТ на востоке США полностью исчез сапсан. Птицы наиболее уязвимы, потому что ДДТ вызывает гормональные изменения, влияющие на обмен кальция. В результате скорлупа откладываемых яиц становится тоньше и они значительно чаще разбиваются. Содержание ДДТ в жировых тканях человеческого тела достигает в США 12–16 частей на млн., тогда как официально установленный предел допустимого содержания его в пищевых продуктах составляет 7 частей на млн.
Позже был создан ряд сильнодействующих, но быстро разрушающихся пестицидов, таких как органофосфаты (например, малатион), и применение ДДТ резко сократилось. Однако производство ДДТ обходится сравнительно дешево, и он по-прежнему остается наиболее пригодным для определенных целей, например, для борьбы с малярией. Решая вопрос о применении пестицидов, мы часто вынуждены из двух зол выбирать меньшее. Благодаря ДДТ во многих районах земного шара удалось полностью искоренить малярию.
5.3. БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ (КРУГОВОРОТЫ)
Подход к познанию экосистем состоит в исследовании больших биогеохимических циклов (круговоротов), различные фазы которых протекают внутри разных экосистем. Речь идет о циркуляционном движении химических элементов абиотического происхождения, которые характерными для них путями попадают из окружающей среды в организмы и из организмов в окружающую среду. Минеральные элементы проникают в ткани растений и животных в процессе их роста и там входят в состав органических веществ; когда же после смерти организма эти элементы вновь попадают в окружающую среду, они перераспределяются, что сопровождается сложными трансформациями и транслокациями, лишь после этого они попадают в новые организмы.
К главным циклам относят биогеохимические циклы углерода, воды, азота, фосфора, серы, биогенных катионов.
Эти циклы не изучаются во всем их объеме, т. е. в масштабе биосферы; их отдельные частные фазы нередко ускользают от нашего внимания, т. к. протекают внутри малоизученных экосистем и к тому же образуют в них вторичные циклы. Иногда эти вторичные циклы связывают несколько экосистем, обеспечивая тем самым максимальное объединение той огромной системы, которую мы зовем биосферой.
Круговорот углерода. Источники углерода в природе столь же многочисленны, сколь и разнообразны. Между тем только углекислота, находящаяся либо в газообразном состоянии в атмосфере, либо в растворенном состоянии в воде, представляет собой тот источник углерода, который служит основой для переработки его в органическое вещество живых существ. захваченная растениями, эта углекислота в процессе фотосинтеза превращается в сахар:
(nCO2 + mH2O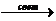 Cn(H2O)m + nO2),
Cn(H2O)m + nO2),
а другими процессами биосинтеза преобразуется в протеиды, липиды и т. д. Эти различные вещества служат углеводным питанием для животных. С другой стороны, все организмы дышат и выбрасывают в атмосферу углерод в форме углекислоты. Когда же наступает смерть, то сапрофаги и биоредуценты разлагают и минерализируют трупы, образуя цепи питания, в конце которых углерод нередко вновь поступает в круговорот в форме углекислоты («почвенное дыхание»).
В известных условиях накапливающиеся мертвые растительные и животные остатки замедляют круговорот углерода: животные-сапрофаги и сапрофические микроорганизмы, обитающие в почве, превращают накопившиеся на ее поверхности остатки в новое образование органической материи, более или менее мощный слой коричневой или черной массы – гумус. Скорость воздействия разлагающихся организмов на гумус далеко не одинакова, а цепи грибов и бактерий, приводящие к окончательной минерализации углерода, бывают различной длины; вследствие этого гумус разлагается то более или менее медленно, то более или менее быстро.
Иногда цепь бывает короткой и неполной: цепь сапрофагов лишается возможности функционировать из-за недостатка воздуха или вследствие слишком высокой кислотности; органические остатки накапливаются в форме торфа и образуют торфяные болота. В некоторых торфяных болотах с пышным покровом из меха слой торфа достигает мощности 20 м и более. Здесь и приостанавливается круговорот. Скопления ископаемых органических соединений в виде каменного угля и нефти свидетельствуют о стагнации круговорота в масштабах геологического времени.
В воде также происходит стагнация круговорота углерода, т. к. углекислота накапливается в форме CaCO3 (мел, известняки или кораллы) химического или биогенного происхождения. Часто эти массы углерода оставались вне круговорота в течение целых геологических периодов, пока CaCO3 в виде горных цепей не поднимался над поверхностью моря. С этого момента начиналось поступление углерода и кальция в круговорот. Оно осуществлялось вследствие выщелачивания известняка атмосферными осадками или под действием лишайников, а также корней цветковых растений. Углерод, накопившийся в почве или в горных породах, может быть освобожден и в процессах человеческой деятельности: горение (отопление, промышленность), обжиг извести.
Круговорот азота. Несмотря на величайшую сложность, этот круговорот осуществляется беспрепятственно и быстро.
Воздух, содержащий 80 % азота, одновременно служит и огромным вместилищем и предохранительным клапаном системы, он беспрерывно и в разных формах питает круговорот азота. Кроме того, электрические разряды, сопровождающие грозы, синтезируют из атмосферного азота и кислорода оксиды азота: эти оксиды попадают в почву вместе с дождевыми водами. Таким путем в экосистеме в форме селитры или азотной кислоты накапливается от 4 до 10 кг азота на 1 га в год.
Происходит и фотохимическая фиксация азота.
Однако наибольшие количества этого элемента поступают в экосистему в результате деятельности микроорганизмов – фиксаторов азота. Чаще всего эту функцию осуществляют бактерии, способные использовать энергию дыхания для прямого усвоения атмосферного азота и синтезирования протеидов. Иногда эти бактерии – аэробы (Azotobacter) или анаэробы (Clostidium) – действуют изолированно; их трупы обогащают почву органическим азотом, который быстро минерализуется. Таким путем в почву ежегодно вносится еще около 25 кг азота на 1 га.
Наиболее эффективны бактерии, живущие в симбиозе с бобовыми растениями в клубеньках, развивающихся на корнях этих растений. А в присутствии молибдена, который служит катализатором, и особой формы гемоглобина (уникальный случай у растений) эти бактерии (Rhizobium) ассимилируют громадные количества молекулярного азота. Образующийся органический азот постоянно диффундирует в ризосферу (ту часть почвы, которая соприкасается с корнями), в особенности когда клубеньки по мере старения распадаются; кроме того, азот в значительном количестве проникает в наземные органы растения-хозяина. Благодаря этому бобовые исключительно богаты протеидами и очень питательны для травоядных животных. Годовой запас, накапливаемый в наземных и подземных органах этих растений, достигает в культурах клевера и люцерны от 150 до 400 кг/га.
Таким образом, существует возможность поддерживать урожайность полей как введением севооборотов, содержащих бобовые, так и использованием азотных удобрений. Современная агротехника рекомендует вносить под посевы бобовых культур особенно активные формы бактерий из рода Rhizobium.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 |
