
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Для амфибий с биологической точки зрения очевидна важность различения в первую очередь движущихся объектов. При этом нейроны, реагировавшие на движение малых объектов, могли не отвечать на движение больших (R. M. Gaze, M. J. Keating, 1970). И, кроме того, нейроны поверхностной части КСМ не изменяли своей активности при сдвигах общей освещенности, в то время как в глубоких слоях отмечались on - и off-реакции. Что касается дирекциональной избирательности, то она, задаваясь отчасти на уровне сетчатки, обычно не только воспроизводится центральными нейронами, но и оказывается здесь более острой, чем у нейронов сетчатки. Такие отношения описаны для НКТ кролика Левик (W. R. Levick et al., 1969). По модели Левик обострение избирательности может происходить за счет конвергенции на каждом нейроне НКТ возбудительных и тормозных входов от нескольких дирекционально избирательных ГКС (по крайней мере, от двух) с оптимальными направлениями под углом 180° между ними.
У амфибий степень избирательности к скорости (O.-J. Grusser et al., 1969) и направлению движения (J. Y. Lettvin et al., 1959) в сетчатке невелика. Одни и те же ГКС реагируют на оба параметра движения. Поэтому у лягушки функция вычисления скорости движения стимула и уточнение направления его движения осуществляется нейронами КСМ, к которым доставляется первичная информация ( и др., 1976; и др., 1977). В поверхностных слоях КСМ локализованы элементы, преимущественно лишь констатирующие наличие движения без определения его направления и скорости, а в более глубоких структурах происходит обострение дирекциональной и скоростной избирательности. Большое значение при этом имеет механизм латерального торможения.
Наличие в реакциях тектальных нейронов, связанных с детекцией параметров движения объекта, стереотипно повторяющихся паттернов импульсов дало основание предполагать, что существенным механизмом вычисления параметров движения является их когерентное накопление. Поэтому детекция движения может эффективно осуществляться нейронами с фоновой активностью, которые способны одновременно участвовать в нескольких независимых процессах обработки сигналов (, , 1975).
Реакция нейронов, не строго специализированных в отношении параметров движения, характеризуется изменением частоты импульсации при изменении скорости или направления движения (D. Finkelstein et al., 1965; O.-J. Grusser et al., 1968, 1969; , 1970; и др., 1977) и изменением ЛП реакции ( и др., 1977; , 1980). В высших отделах мозга птиц и млекопитающих описаны подобные реакции нейронов, связанных с детекцией направления и скорости движения зрительного стимула-объекта. Так, D. Jassik-Gershenfeld et al., (1970) показали, что 35% нейронов tectum opticum голубя, реагировавших на движение, обладали избирательной чувствительностью к направлению движения. У кролика предпочитаемые направления для всех исследованных детекторов коры распределились в четырех неперекрывающихся группах вдоль вертикальной и горизонтальной осей поля зрения (C. W. Oyster, H. B. Barlow, 1967), Интересно, что предпочитаемые направления для детекторов сетчатки рыбы также распределились только по четырем направлениям вдоль вертикальной и горизонтальной осей поля зрения, причем клетки, чувствительные к горизонтальному направлению, встречались чаще (I. K. Cronly-Dillon, 1964). Нейроны же тектума лягушки реагируют значительно сильнее на движение стимула вверх-вниз, чем вправо-влево (K. V, Fite, 1969; и др., 1976, 1981), что, видимо, находится в связи с большим развитием вертикальных движений глаз у лягушки во время дыхания, чем горизонтальных. Скорость движения наряду с направлением является важным фактором, определяющим реакции большинства нейронов тектума голубя (, 1970); наибольшая степень избирательности обнаружена у клеток средних и глубоких слоев. На уровне коры у кролика показано, что большинство клеток (56%) чувствительны к движению стимула, при этом 20% корковых нейронов обладает избирательностью к направлению движения (И. Кетлерис, 1979); среди них обнаружены как моно - (71,4%), так и бидирекционально избирательные элементы (28,6%). По данным И. Кетлериса (1979), в зрительной коре кролика представлены в различном количественном соотношении все возможные (от 0 до 360°) оптимальные направления, однако 48% дирекционально избирательных клеток предпочитали направление движения стимула параллельно вертикальному и горизонтальному меридианам поля зрения, причем у этих нейронов степень дирекциональной настройки оказалась значительно выше, чем у клеток с диагональными оптимальными направлениями. И при этом проявление дирекциональной избирательности оказалось зависящим от скорости движения стим9; и др., 1981). В верхнем двухолмии млекопитающих описаны нейроны, селективные к скорости движения стимула (Y.-C. Tiao, C. Blakemore, 1976): среди нейронов поверхностных слоев с РП 10° 80% имели симметричную структуру и реагировали на медленно движущиеся стимулы, а 6% нейронов отвечали оптимальную скорость 200°/с; остальные нейроны (14%) проявляли дирекциональную избирательность. В средних и глубоких слоях верхнего двухолмия размер РП увеличивался. У многих нейронов наблюдался эффект привыкания. Однако собственно детекторов скорости на уровне верхнего двухолмия кролика не описано. В верхнем двухолмии кролика не наблюдается острой избирательности к узкому диапазону скоростей, как это описано дл нейронов сетчатки кролика (H. B. Barloy et al., 1964), для коры и верхнего двухолмия белки ( и др., 1973). Часть нейронов верхнего двухолмия кролика не обнаруживает заметной избирательности к скорости движения стимула, хотя у большинства нейронов этой структуры такая зависимость имеет место. Предполагается, что важная роль в формировании чувствительности нейронов к малым и большим скоростям принадлежит пространственно-временной суммации в РП нейронов возбуждения и торможения (, 1974). В верхнем двухолмии высших млекопитающих (обезьян) описаны нейроны, которые давали ответы на включение и выключение неподвижного светового пятна и на движущееся изображение, причем они реагировали на движение независимо от его направления, но меняли реакцию при изменении скорости движения (M. E. Goldberg, R. H. Wurtz, 1972). Дирекциональная избирательность проявляется и у нейронов НКТ (G. B. Arben, 1962, 1963; W. Kozak et al., 1965; W. R. Levic et al., 1969), а избирательности к скорости движения на уровне НКТ не описано. В зрительной коре, как и на других уровнях зрительного анализатора, избирательность сенсорных нейронов к направлению скорости движения зрительного стим4, 1981).
По чувствительности к скорости движения зрительного стимула нейроны можно разделить на индифферентные и выделяющие определенные скорости и их диапазоны (G. Orban, 1975; С. Алексеенко и, 1976; и др., 1977; , 1980). Среди них есть нейроны, проявляющие избирательную чувствительность к определенной скорости движения стимула. И. Кетлерисом (1979) на зрительной коре кролика и и др. (1977, 1982) на тектуме лягушки показано, что возможны два вида реакций нейронов на скорость движения: узкая настройка нейрона на небольшой диапазон скоростей (собственно детекторы скорости) и работа нейрона широкополосного фильтра, меняющего свой ответ в зависимости скорости движения; при этом оказалось, что узкополосные нейроны формируются из широкополосных на более высоком уровне системы. Поскольку оптимальная скорость для разных нейронов различна, то возможно, что совокупность нейронов, чувствительных к разным скоростям движения, образует систему, классифицирующую стимулы по, скорости их движения (, 1974; и др.; 1977; , 1980). Видимо, можно предположить что избирательность к определенной скорости движения может кодироваться номером канала, т. е. тем или иным нейроном (; 1974; И. Кетлерис, А. Пуртулите, 1976; и др., 1977) Для лягушки в ГКС кодирование скорости движения осуществляется только числом импульсов в ответе (D. Finkelstein, O.-J. Grusser, 1965; O.-J. Grusser et al., 1968; , 1970), и поэтому детектирование скорости можно предполагать лишь на высших уровнях зрительной системы. В зрительной системе амфибий и в зрительной системе грызунов среди нейронов, реагирующих на движение стимулов, были найдены детекторы малой и большой скорости. Механизмы, обеспечивающие такую избирательность нейронов, определяются соотношением тормозных процессов и эффективностью пространственно-временной суммации возбуждения, вызываемого воздействием движущегося стим2, 1973). Показано, что нейроны с простым РП чувствительны к низким скоростям движения, а со сложными - к более высоким (L. Movshon, 1975). В реакциях нейронов, избирательных к высоким скоростям движения стимула, в зрительной коре кролика четко выделяются фазические и тонические компоненты. Возможно, такое разделение реакции на компоненты может отражать поэтапную реализацию функций обнаружения и опознания объекта (, 1974). По мнению ряда авторов (И. Кетлерис и др., 1973; и др., 1975, 1976, 1981), конечным этапом анализа движения объекта является синтез скорости и направления движения, осуществляемый нейронами-интеграторами, кодирующими одновременно оба параметра (скорость и направление движения), однако в психофизиологических опытах получены результаты, указывающие на наличие раздельных систем анализа направления и скорости движения (B. Breitmeyer, 1973).
1.2.1. Организация рецептивных полей тектальных нейронов
Исследования РП ретинальных (H. B. Barloy et al., 1957; J. Y. Lettvin et al., 1959, 1961; H. B. Barloy, W. R. Levick, 1965; , 1977; , 1980), тектальных (,1969; и др., 1976; и др., 1977, 1982) таламических (O. H. Hubel, T. N. Wiesel, 1961; и др., 1973) и корковых (O. H. Hubel, T. N. Wiesel, 1959, 1965, 1968; и др., 1972, 1973, 1977; , 1972, 1973, 1975, 1978, 1981; и др., 1974, 1979, 1982; Л. Митова, 1977) нейронов показали, что наряду с концентрическими РП в зрительной системе достаточно большое место занимают вытянутые и асимметричные поля, которые обеспечивают выделение таких параметров объекта, как его ориентация (L. A. Movshon, 1979; , 1979), направление движения (H. B. Barloy, W. R. Levic 1965; и др., 1976; , 1979; и др., 1981) и некоторые признаки формы (J. V. Lettvin et al., 1959). Хотя для лягушки характерен детекторный принцип опознавания уже на ретинальном уровне, однако в ее ретине описаны и концентрические РП (J. V. Lettvin et al., 1959, 1961; , 1980). Что касается следующего этажа обработки зрительной информации лягушки - tectum opticum, то имеющиеся отдельные исследовав РП нейронов этого уровня не позволяют составить более или менее систематизированное представление об их форме. Поэтому нами было проведено специальное исследование РП тектальных нейронов. Для этого в одной серии опытов все поле зрения лягушки сканировалось световым пятном с диаметром 3° (если на него не было реакции, то оно заменялось пятном с диаметром 10° и т. д.), а в другой серии через поле зрения в 8 направлениях (под углом друг к другу в 45°; перемещался стимул-объект (темный прямоугольник с угловыми размерами 0,5x2,6°, движущийся вдоль своей длинной стороны со скоростью 18°/с). Для оценки наличия реакции строились пре - и постстимульные гистограммы и достоверность ее оценивалась критериям Стьюдента и Фишера при β≥0,95.
В первой серии опытов со стационарным стимулом в случае обнаружения четких границ поля в центр его последовательно проецировались пятна с диаметрами 3, 10 и 30° (П3°, П10°, П30°), а также использовался ДЗ. Пятна предъявлялись глазу лягушки на 5 с и исследовалась реакция тектального нейрона на появление (включение) и исчезновение (выключение) пятна, как и на включение выключение ДЗ. Во второй серии для определения примерных размеров и формы РП производился анализ гистограмм, построенных по их активности, проявляющейся при движении стимула в разных направлениях.
При исследовании РП тектальных нейронов стационарными вспыхивающими пятнами и ДЗ обнаружилось, что в ростральной и каудальной частях тектума соотношение возбудительных и тормозных реакций на разные стимулы имеет существенные различия лишь при анализе мелких стимулов (П3°) и перекрывающих поле зрения (ДЗ). Для суждения о гомо - или гетерогенности поля исследовался знак реакции нейрона на возрастающую по площади стимуляцию, а также “выраженность” реакции нейрона, за которую условно принимали разность между средней частотой во время реакции и с новой частотой импульсации нейрона (для Т-реакций фоновоактивных нейронов эта величина была отрицательной). При этом повышение “выраженности” ответа при сохранении знака реакции на увеличивающуюся по площади стимуляцию говорило о суммаци возбуждения (или торможения) в гомогенном РП, а снижение-- гетерогенности поля, возможно, о перекрытии В - и Т-зон, о диффузном представительстве в зоне, снижающей ответ, реципрокности строенных единиц, суммарное возбуждение которых (при доминировании одного из процессов) приводит к уменьшению ответа, хотя знак реакции, навязанный доминирующими единицами, не инвертируется, а сохраняется прежним. Оказалось, что только 9% исследованных РП являлись гомогенными, в которых происходили процессы пространственной суммации. В этих случаях размер стимула мог кодироваться интенсивностью реакции - с помощью частотных составляющих кода, длины пачки импульсов, разностей ЛП ответов и т. д. В некоторых случаях отмечался “волнообразный” характер реагирования нейрона при изменении стимулов от малого пятна до ДЗ: сначала увеличение ответа, потом. уменьшение его, а затем снова увеличение, т. е. в этих случаях РП имело “слоистую” структуру: в центре возбуждение, вокруг торможение и снова возбуждение (или наоборот для Т-центральных полей). Таких гетерогенных “слоистых” полей оказалось около половины всех гетерогенных (и почти 27% от числа всех реагирующих). Таким образом, предъявление стимулов разных размеров выявило перекрытие облегчающих и тормозных зон в РП тектальных нейронов. Похожие отношения при предъявлении полос были получены на корковых нейронах кошки Митовой (L. Mitova, 1976; Л. Митова, 1977).
В опытах со стимулом, движущимся в разных направлениях выявились РП весьма разнообразной формы с изолированными и перекрывающимися on - и off-зонами, что и определяет в значительной степени их дирекциональную избирательность. Так, около 23% из всех реагирующих нейронов имели различные формы круглых полей, около 57% имели вытянутые РП различной формы и около 20% - поля, которые, являясь круглыми или вытянутыми, представляли собой довольно своеобразные картины из сочетаний on - и off-зон, часто организованных мозаично.
Среди круглых полей основная масса (63% из количества всех круглых полей) характеризовалась гетерогенностью зон, причем некоторые из них имели разделенные В - и Т-зоны, а некоторые-частично перекрывающиеся. Кроме того, имели место и гомогенные В - и Т-РП округлой формы, составившие меньшую количественную группу - 37% (рис. 2). При этом диаметр круглых РП достигал 30÷42° при диаметре центральной зоны от 2÷3° и 6÷8° до 12÷18°.
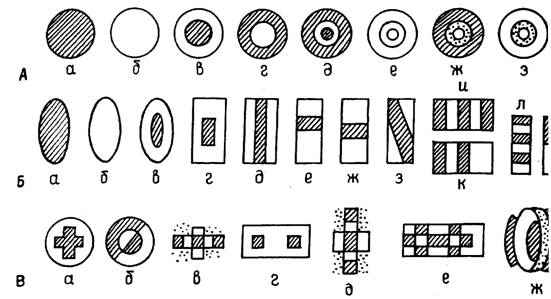
Рис. 2. Различные виды круглых (А), вытянутых (Б) и "мозаичных" (В) РП тектальных нейронов лягушки; заштрихованные участки - В-зоны; светлые - Т-зоны; пунктирные - зоны, в которых реакция отсутствует.
Вытянутые поля также были представлены и гомогенными В - и Т - РП (13% от всех вытянутых полей), и РП, разделенными на гетерогенные зоны (87%), среди которых также были РП с частичным перекрытием В- и Т-зон (см. рис. 2). Длина вытянутых полей колебалась от 30 до 42°, а ширина - от 2÷6° до 12÷18° при длине центральной зоны от 6 до 18° и ширине от 2 до 6°. Следует специально отметить структуры вытянутых полей с чередованием полос возбуждения и торможения: ширина таких полос составляет 6-8°, а длина от 12÷18° до длины РП (30÷42°), когда эти полосы, чередующиеся по знаку реакции (В, Т, В, Т, и т. д.), вытянуты вдоль РП во всю его длину. Такие “полосатые” РП тектальных нейронов напоминают корковые РП млекопитающих с разделенными on - и off-зонами. Частичное перекрытие on - и off-зон, наблюдаемое в ряде РП лягушки, так имеет место и в корковых полях высших позвоночных.
Поля третьей группы, которые мы условно обозначаем как “мозаичные”, представляют собой весьма разнообразные сочетания В - и Т - зон. Эти РП, являясь производными от круглых и вытянутых полей, имеют примерно такие же размеры: в случае круглых “мозаичных" РП диаметр их был 30÷42° при диаметре центральной зоны от 2 до 18°, а в случае вытянутых “мозаичных” РП их длина составляла 30÷42°, ширина 6÷18° при длине центральной зоны 6÷18° и ширине 2÷6°. Для большинства “мозаичных” РП, включающих несколько В - и Т-зон, вообще трудно говорить о каком-либо центре и периферии, так как эти поля представляют собой чередующиеся зоны возбуждения и торможения, часто в виде довольно сложной мозаики (см. рис. 2).
Большинство тектальных нейронов имеют РП, по-разному организованные в зависимости от того, в каком направлении (имеются в виду встречные направления) движется тестирующий стимул, одна основные описанные нами виды полей при этом сохраняются. В случае дирекционально не избирательных или мало избирательных нейронов выявились более или менее симметричные РП, в то время как дирекционально избирательные нейроны имели РП с более выраженной ориентацией. Для таких нейронов приводятся рисунки РП, снятых при движении стимула в оптимальном направлении и в направлениях, прилежащих к оптимальному. На наш взгляд, следует обратить внимание на “мозаичные” РП, которые имеют довольно упорядоченную конструкцию (не всегда симметричную, что создает условия их дирекциональной неравнозначности) и отличаются малыми В - (а в ряде случаев и малыми Т-) зонами. По-видимому, для таких полей наш стимул (темный “жук” с размерами 0,5x2,6° (является по величине вполне адекватным (наиболее характерные размеры “мозаичных” off-В - и off-Т-зон: 2x2°, 6x6°, 8х8°, 4x6°, 8x16° и т. д.). Естественно, наиболее адекватные условия стимуляции достигались при движении оптимально ориентированного стимула в оптимальном направлении с оптимальной скоростью. Чаще всего это была вертикальная ориентация при движении в вертикальном направлении сверху вниз со скоростью 18°/с (K. Fite, 1969; и др., 1976; и др., 1977, 1981). Однако для ряда нейронов это были другие значения ориентации, направления и скорости.
Исследование организации РП тектальных нейронов на разной глубине КСМ выявило некоторую зависимость структуры РП от локализации нейронов в тектуме: нейроны поверхностных слоев имеют РП, организация которых весьма сходна с таковой у ГКС ( и др., 1982). Это, вероятно, в значительной степени связано с тем, что здесь локализованы собственные ретинальные волокна, имеющие часто гомогенные круглые и вытянутые РП и такие же по форме поля, но концентрические с гетерогенными зонами (глубина 100÷300 мкм). Однако и на значительной глубине (до 500÷700 мкм) встречаются такие же поля, хотя здесь чаще локализован нейроны с “полосатыми” и “мозаичными” РП. Наличие таких тектальных нейронов с “полосатыми” и “мозаичными” РП говорит сложной конвергенции на их мембране путей от ГКС, а возможно, от ряда тектальных нейронов с круглыми и вытянутыми гомогенным и концентрическими РП. Такая множественная конвергенция пут от ретинальных и тектальных нейронов при сложной и разнообразной организации их возбудительных и тормозных синаптических мест на мембране адресного нейрона и в зависимости от их синаптических весов приводит к возникновению различных конструкций “мозаичных” РП, которые, по-видимому, выполняют интегративную функцию, являясь специфическими анализаторами движущихся мелких зрительных объектов.
1.2.2. О вероятности в характере реакций нейронов тектума
Одной из характерных черт деятельности нейронов высших отделов мозга является вероятностный способ их функционирования (А. Фессар, 1962; , , 1965; , 1972), что отражает особый характер деятельности, войственный нейронам информационно-управляющих структур (D. Hebb, 1949; Б. Бернс, 1968), и обеспечивает высокую надежность и полифункциональность нейронных систем мозга. Возможно, и нейроны, осуществляющие детекторные функции и организующиеся при действии оформленного стимула в определенные группировки, также используют вероятностный принцип взаимодействия ( и др., 1973; , 1974; и др., 1976). В литературе описано “привыкание” к повторным раздражениям нейронов “новизны” и опережающие ответы нейронов “тождества”. Кроме того, (1967) наблюдал явление стабилизации ответов нейронов, которую он рассматривает как характеристику перестройки нейронных сетей и связывает с усилением возбуждения и торможения, для чего служит сеть с латеральным торможением, повышающим контраст восприятия.
Проводившиеся нами исследования реакций детекторных неронов тектума лягушки обнаружили имеющий в ряде случаев место их вероятностный характер, проявляющийся в изменении реагирования нейронов в ходе их “привыкания” к стереотипной стимуляции возникновении экстраполяционных ответов на повторное применение стереотипного стимула. С целью изучения вероятностного реагирования нами были исследованы адаптационные и экстраполяционные характеристики нейронов КСМ лягушки, связанных с детекцией признаков формы и дирекционально-скоростной чувствительностью. В качестве стимула-объекта применялся темный прямоугольник с угловыми размерами 0,3÷1,3х0,6÷2,6° (преимущественно - 0,5х2,6°), движущийся с постоянной скоростью 10÷20°/с (преимущественно - 18°/с) вдоль зеленой полосы линии развертки Преимущественное направление движения - сверху вниз (хотя пользовались и другие направления). Стимул предъявлялся глазу лягушки 10-кратно с интервалами 30 с в одной серии опытов и 5 с - в другой. У многих нейронов при повторной стимуляции отмечалась вероятностность в реагировании, заключавшаяся в довольно значительной вариабельности ЛП ответов, в изменении длительности выраженности реакции, а в ряде случаев и ее знака (переход Т-реакций в В-реакции и наоборот), в периодическом исчезновении и появлении реакций, в полном прекращении реагирования, в явлении опережающих ответов, а также в изменении характер детектирования. Так, оказалось, что некоторые нейроны, связанные с детекцией направления движения, при повторных пробах могут включаться в ансамбли, участвующие в детекции движущегося края, и наоборот, либо нейроны, дирекционально избирательные к одному направлению, переключают свою избирательность на другое направление и т. д. Отмечавшееся привыкание нейронов к повторному воздействию раздражителей выражалось удлинением ЛП реакции, уменьшением длительности и выраженности ответов вплоть до полного прекращения реагирования. В ряде случаев при стереотипном действии раздражителя наблюдается обратный эффект - укорочение ЛП и появление “экстраполяционных” опережающих раздражение ответов, что, видимо, связано с реализацией “памяти” нейрона. При этом, если раздражитель предъявлялся с интервалом в 30 с, некоторые нейроны (7%) в течение достаточно длительного времени (5-10 мин) имели стабильные параметры реакций (которые могут рассматриваться как кандидаты в коды), но большинство (93%) обладало нестабильными реакциями со значительными флуктуациями ЛП и длительностей ответов, причем лишь немногие из них оказались настоящими нейронами “новизны”, т. е. исключительно “привыкающими” (11%), или нейронами “тождества”, “запоминающими” стимул с опережением в реакции (7%), а основная масса (75%) отличалась вариабельным характером реагирования, обнаруживая в своей деятельности как способность к “привыканию” и к восстановлению реакций (49%), так и к временной стабилизации ответов (13%) и к проявлению “памяти” с “экстраполяцией” и с восстановлением реакций (42%), т. е. основная группа нейронов реагировала настолько вероятностно, что в ответах у одних и тех же нейронов отмечалась способность и к привыканию, и к опережению стимуляции, и к восстановлению первоначального характера реакций.
Если же раздражитель применялся более часто (с интервалами 5 с), только 4% нейронов давали стабильные реакции; остальные 96% нейронов обладали вероятностным типом реагирования: это были и “чистые” нейроны “новизны” и “тождества”, и нейроны, в ответах которых имели место компоненты и привыкания, и экстраполяции, и восстановления с некоторой стабилизацией реакций. По-видимому, нейроны с вариабельными временными характеристиками ответов могут кодировать информацию о параметрах сигнала позиционно номером канала, а внутри канала, если имеются закономерности в изменении вероятности реакции, может быть использовано вероятностное кодирование (Р-кодирование - D. H. Perkel, T. H. Bullock, 1968). Закономерный же сдвиг реакции по шкале времени у нейронов “новизны” и “тождества” позволяет, видимо, нейронам мозга “вычислить” повторность действия стимула при наличии стабильно реагирующих нейронов, создающих “точку отсчета” для вычисляющих систем мозга.
Одной из основных задач современной нейрофизиологии является выяснение закономерностей нейронной организации рабочих механизмов мозга. При этом весьма существенным вопросом является определение места, занимаемого нейроном в пространстве и во времени в системе реагирования. Для выяснения возможной взаимосвязи реакций разных нейронов можно использовать ЛП их ответов (, 1965). В литературе встречаются разные сведения о степени устойчивости ЛП нейронных реакций (O.-J. Grusser, A. V. Grutzner, 1958; , 1965; , , 1973; М. -Гохари, 1974; , 1974; и др., 1976; , 1980). Флуктуации ЛП нейронных реакций нижнего этажа (в частности, в сетчатке) создают предпосылки для вариабельности ЛП ответов тектальных нейронов, которая связана, кроме того, с колебаниями возбудимости самих центральных нейронов. Отсюда, естествен вытекает разная вероятность вовлечения нейронов относительно однородной популяции, одного ансамбля, одной функциональной системы в ту или иную реакцию (, 1969, 1972).
Демонстративным показателем такого временного группирования нейронов, выполняющих общую функцию, являются “цепочки возбуждения” (, 1965), построенные по их ЛП и позволяющие судить о возможном (стабильном или вариабельном) участии нейрона в реакции и его месте в процессе реагирования зрительных ансамблей на специфическую стимуляцию. Для нейронов, реагирующих на движение, исследовались относительные (и условные) ЛП, т. е. время от начала движения стимула от края экрана к его центру до начала возбуждения нейрона. Было показано, что в то время, как засветы вызывают сначала более и менее обширную активацию, которая затем сменяется процессе распространяющимся по более узкому пути, движение в поле зрения оформленного стимула приводит к активации более или менее постоянного количества нейронов в течение довольно длительного времени, что, видимо, связано с постепенным вовлечением при движении стимула разных РП ГКС и соответственно разных тектальных нейронов.
Исследование локализации коротко - и длиннолатентных on-off-ответов по глубине тектума лягушки показало их равномерное распределение по тектальным структурам ( и др., 1976; , 1980; , , 1981). Таким образом, оказалось что относительно коротколатентные нейроны могут локализоваться в более глубоких слоях, чем иные длиннолатентные, и поэтому ход возбуждения может оказаться весьма сложным: сначала, например, могут возбудиться некоторые элементы, лежащие на глубине 500÷600 мкм, а потом - на глуби 100÷300 мкм мкм. Это, видимо, связано как с ветвлением аксонов ГКС на значительной глубине, так и с тем, что в тектуме имеют место восходящие коллатерали, направляющиеся из глубоких слоев в поверхностные (, 1948; G. Szekely, 1973). Регистрация длиннолатентных ответов у нейронов поверхностных слоев может также говорить в пользу ветвления процесса на этом уровне и циркуляции его по клеточным элементам КСМ. Это в равной мере относится как к нейронам, реагирующим на засветы, так и к нейронам связанным с детекторной функцией. Однако в отношении постоянства места нейрона в системе реагирования на засвет и на движущийся стимул такого однообразия нет: видимо, ансамбли нейронов формирующихся при вспышках света разной интенсивности и крутизны, хотя и различаются между собой, но имеют общие черты организации и характеризуются большей стабильностью, чем ансамбли, формирующиеся для опознания более сложного зрительного образа, в частности движущегося с разными скоростями и в разных направлениях объекта. При этом чем проще и постояннее сигнал (вспышка), тем менее вариабельна на него реакция, а чем образ сложнее, тем больше вероятностных элементов в реагировании нейронов.
Изучение характера В-реакций тектальных нейронов в ответ на зрительную стимуляцию приводит к мысли, что ряд характерные компонентов в вызванных раздражением импульсных последовательностях можно рассматривать как кандидаты в коды (D. H. Perkel, T. H. Bullock, 1968). Так, в ряде случаев и on-, и off-, и on-off-нейроны имели стабильные ответы в виде одного-двух импульсов (при отсутствии фоновой активности) либо отмечалось учащение импульсации (при наличии фоновой активности), причем при анализе реакций часто удавалось выделить в ответе нейрона также всего один-два “информационных” импульса, “инъецированных” в фоновую активность. В отдельных случаях имели место высокочастотные пачечные ответы. Наиболее характерным константным компонентом многих реакций являются ЛП ответов или отдельных импульсов в них при неизменной стереотипной стимуляции (хотя часто имеют место и реакции с флуктуирующим ЛП). Следует отметить, что для большинства on- off-нейронов характерны различающиеся между собой on - и off-ответы либо по числу импульсов, либо по длине пачки, либо по паттернам, либо по иным параметрам и почти всегда по ЛП.
Что касается В-реакций нейронов на движение стимула, то у большинства из них отмечалось учащение фоновой импульсации (или появление ее у фоновомолчащих нейронов) при пересечении стимулом РП нейрона. При этом наряду с ответами, характеризующимися стабильной латенцией, отмечались реакции с вариабельными ЛП, а среди них имели место как стабилизирующиеся ответы, так и ответы, угасающие (при привыкании) и усиливающиеся (с экстраполяцией). Часть нейронов отвечала 1-2 импульсами (реже 3) с постоянным ЛП, а часть - постоянным паттерном ( в ряде случаев с нестабильным ЛП первого импульса).
1.2.3. Детекторная функция тектальных нейронов
Для исследования детекторной функции тектальных нейронов мы использовали модифицированную методику Леттвина с соавт. (J. V. Lettvin et al., 1959) и перемещали перед глазом лягушки по внутренней поверхности полусферы, удаленной на 15 от глаза, либо с помощью магнита, либо с помощью специальной дуги различные мелкие объекты: чаще всего черный ромб с диагоналями 13 и 20°, движущийся вперед острым углом и способный таким образом, раздражать РП от долей градуса до 13° и, следовательно, вовлекать в реакцию нейроны, связанные с ретинальными ДВК и ДДК. Было установлено, что 70% нейронов тектума у лягушки связаны с детекцией движения. Из всех детекторных не нов около 36% составили единицы, связанные с ДДК, около 44% - с ДНД и примерно 20% - с ДВК. Некоторые нейроны лишь условно могли быть отнесены к одной из групп, так как они реагировали либо меняя характер детектирования, либо имея характеристики ответа, напоминающие разные виды детекции. Так, некоторые нейроны, отвечающие на стимулы средней величины (как ДДК), в то же время сохраняли реакцию при остановке объекта в их РП (как ДВК), что обусловлено, видимо, конвергенцией на этих нейронах путей, проводящих возбуждение от соответствующих ретинальных детекторов или связанных с ними тектальных нейронов. Анализ распределения собственно тектальных элементов, связанных с детекцией разных признаков, показал, что тектальные нейроны, сходные по своим свойствам с ретинальными детекторами, распределены по всем тектальным структурам равномерно.
При наличии обоих типов реакций (В и Т) чаще встречались нейроны с тормозным характером реагирования на движение стимула-объекта в РП. Так, около 60% реагирующих нейронов отвечали только Т-реакциями, 30% - только В-реакциями и 10% - В - и Т-реакциями. Подобное преобладание Т-ответов описано для нейронов tectum opticum при световой стимуляции и при раздражении зрительного нерва (, 1969; , А С. Лукьянов, 1971; , 1973; , 1974; , 1980; , 1980). Торможение тектальных нейронов может быть следствием как центрального возбуждения соседних нейронов КСМ, так и торможения нйронов сетчатки, т. е. это либо латеральное (и, возможно, возвратное) торможение тектальных нейронов, либо результат латерального торможения ретинальных нейронов. Оно является важным механизм выделения признака объекта и должно играть существенную роль в детекторной функции (H. K. Hartline, 1949; H. B. Barloy, 1953, , 1966). При анализе распределения В - и Т-ответов нейронов по глубине тектума также было обнаружено равномерное их распределение по тектальным структурам. При этом почти во всех слоях наблюдалось преобладание Т-реакций.
Для нейронов зрительной системы лягушки ключевыми раздражителями, по-видимому, являются параметры движения стимул (направление и скорость) и размеры его, так как именно эти признаки должны обусловить реакцию животного “на добычу” или “на врага”. Поэтому и нас привлекали для изучения организации зрительных нейронов в структурах мозга лягушки именно эти признаки стимулов. В качестве раздражителей разного размера использовались световые пятна с диаметрами 3, 10 и 30°, вспыхивающие н экране и гаснущие, а также тотальный ДЗ, а для исследования чувствительности нейронов к изменению направления движения скорости использовался движущийся стимул-объект с угловыми раз мерами 0,5х2,6°, способный перемещаться в 8 направлениях несколькими фиксированными скоростями (от 6 до 60°/с).
Оказалось, что при включении светового пятна диаметром 3 (П3°) в каудальной части тектума количество В - и Т-ответов существенно не различается, в то время как в ростральной тектальной зоне выявляется значительно больше тормозных реакций, чем воз будительных. Такие же отношения имеют место при выключении П3°. Включение и выключение П10° и П30° не выявило существенных отличий в количестве В - и Т-реакций ни в ростральной, ни в каудальной зонах тектума. В то же время ДЗ вновь обнаружил неоднородность в реагировании нейронов передней и задней половин тектума: так, в каудальной части тектума и включение и выключение ДЗ вызывают существенно больше Т-реакций, чем В-, при отсутствии таких различий в ростральной части.
Что касается характера реакций нейронов тектума на включение света, то треть on-нейронов, дававших В-ответы, реагировала одним импульсом со стабильным ЛП, у части нейронов в ответе возникало 1-2 импульса, причем вероятность появления второго импульса зависела от наличия первого (при отсутствии первого чаще появлялся второй), что, возможно, позволяет считать этот второй импульс “дублирующим”. У ряда нейронов отмечался пачечный ответ с постоянным ЛП для одного из импульсов или с характерным паттерном, отличающимся стабильными межимпульсными интервалами. И, наконец, часть нейронов отвечала на включение света появлением (или исчезновением) активности либо увеличением (или уменьшением) ее частоты и длительности.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
