
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Аналогичные результаты дало использование в качестве раздражителя вспышки, в ответ на включение которой также возникали реакции одиночными импульсами или пачечные разряды с постоянным ЛП одного или нескольких импульсов. Однако эти ответы, обладая постоянными компонентами при действии данного раздражителя, отличались от ответов этих же нейронов на включение и выключение ДЗ. При действии более яркой вспышки отмечалось закономерное укорочение ЛП ответов, часто перестройка паттернов с укорочением ЛП “информационного” импульса, а также у ряда нейронов имело место увеличение частоты ответа (нередко с укорочением пачки) либо удлинение пачки (без учащения), т. е. использовалось кодирование числом импульсов, В ряде случаев при стереотипной стимуляции отмечалось изменение вероятности ответа (Р-кодирование).
Следует особо остановиться на кодировании освещенности разностью ЛП ответов тектальных нейронов (аналогично описанному для ГКС). Исследование реакций тектальных нейронов показало, что наряду с нейронами, обладающими вариабельными характеристиками ответов (чаще это были фоновоактивные), имели место и такие единицы (чаще фоновомолчащие), которые реагировали на засветы с постоянными ЛП реакций, что говорит о возможн кодирования реакций этих нейронов разностью их ЛП. Исследование степени вариабельности конструкций межимпульсных интервалов в паттернах ответов фоновомолчащих нейронов тектума лягушки на повторную стимуляцию стереотипным световым раздражителем позволило обнаружить относительно постоянный набор межимпульсных интервалов в ответах ряда нейронных единиц.
Таким образом, общая освещенность может кодироваться в тектуме, во-первых, пространственно - on-нейронами, во-вторых, различными видами временных импульсных кодов: разностью ЛП ответов или отдельных импульсов в ответах, изменением частоты пульсации и числа импульсов в пачке, длительностью пачки и перестройкой паттерна и т. д.
На выключение ДЗ больше половины фоновоактивных нейронов с off-ответами реагировало длительным повышением частоты пульсации (до 200-500 мс), а фоновомолчащие нейроны отвечали появлением импульсации, причем часть из них имела стабильный ЛП хотя бы одного (не всегда первого) импульса (частотно-латентный код), а часть нейронов, изменявших частоту импульсации характеризовалась вариабельными ЛП (частотный код). В некоторых случаях наблюдалось повышение (или уменьшение) вероятности ответа при повторных раздражениях, и наступала стабилизация ответа с 3-5-го предъявления стимула - “вероятностное кодирование”. Часть нейронов отвечала на выключение света появлением одного импульса с нестабильным ЛП, что говорит в данном случае об информативности наличия импульса. Часть же нейронов реагировала 1-2 импульсами, один из которых можно, видимо, рассматривать как дублирующий; при этом ЛП хотя бы одного импульса (а чаще обоих) был постоянным, И здесь также следует отметить в качестве “кандидата” в коды разность ЛП off-ответов нейронов.
Таким образом, затемнение, как и освещение, кодируется, с одной стороны, пространственно - off-нейронами, а с другой - степень его выражается временными кодами (латентный, частотный, числовой, паттерновый); определенную роль также играет вероятностное кодирование.
Что касается on-off-нейронов, то здесь не приходится говорить о пространственном кодировании освещения или затемнения, а можно предполагать лишь пространственное кодирование перепада освещенности без учета его знака. Знак же и количественное значение градиента освещенности кодируется в этих нейронах различными видами временных кодов и особенно разностью ЛП on - и off-ответов и различием on - и off-паттернов при постоянстве микроструктуры отдельно внутри on-паттернов и отличающихся от них off-паттернов. Некоторые нейроны отвечали пачечными разрядами на один из компонентов засвета (включение или выключение) и одним-двумя импульсами на другой, зачастую со стабильным ЛП. При этом в большинстве случаев ЛП on-ответа на 30-100 мс оказывается длиннее, чем ЛП off-ответа. У части нейронов ответ на включение и выключение состоял из 1-2 импульсов со стабильными ЛП и, как правило, off-реакции этих нейронов имели более короткие ЛП, чем on-реакции. Часть нейронов on-off-группы и на включение, и на выключение света отвечала учащением импульсации (в случае наличия фоновой активности) или появлением ее (у фоновомолчащих нейронов), при этом ЛП реакции часто был вариабелен. Однако в реакциях отдельных нейронов не удалось выявить специфические компоненты для on- и off-ответов. Видимо, эти нейроны могут лишь констатировать наличие перепада в освещенности без указания знака его.
Таким образом, в группе on-off-нейронов пространственно кодируется лишь наличие градиента освещенности, а знак его и степень перепада (крутизна изменения) уточняется с помощью различных модификаций временных кодов: латентно-частотного, латентно-паттернового и т. д.
Появление и исчезновение в РП стационарных стимулов разной величины также может кодироваться, во-первых, адресно - on-, off - и on-off-нейронами и нейронами, связанными с ДК, ДВК, ДДК и НЗ, и, во-вторых, яркость и размер стимула может уточняться с помощью временных кандидатов в коды (интенсивностью реакции, т. е. частотным и числовым кодами, и изменением ЛП ответов, т. е. кодом разностью ЛП, или Т-кодированием (D. H. Perkel, T. H. Bullock, 1968)).
При движении оформленного стимула в РП тектальных нейронов у последних могут возникать весьма специфичные ответы, характеризующиеся определенным временным узором импульсации. При том наряду с ответами, отличающимися стабильной латенцией отмечались реакции с вариабельными ЛП, а среди них имели место как стабилизирующиеся ответы, так и угасающие (при “привыкании” нейронов “новизны”), и усиливающиеся (часто с экстраполяцией у нейронов “тождества”). Часть нейронов отвечала 1-2 импульсами (реже 3) с постоянным ЛП, а часть - постоянным паттерном (в ряде случаев с нестабильным ЛП ответа). Подобные реакции с постоянством ЛП и относительно устойчивыми паттернами отмечались не только у фоновомолчащих, но и у некоторых фоновоактивных нейронов тектума лягушки в ответ на повторную стимуляцию контралатерального глаза движущимся стимулом, хотя для большинства тектальных нейронов в условиях такой стимуляции было характерно вероятностное реагирование с непостоянным ЛП и различающимся характером импульсации в ответе ( и др., 1976). Однако и при вероятностном реагировании оказалось возможным в ряде случаев выделить в постстимульных последовательностях фоновоактивных тектальных нейронов одиночные импульсы, появляющиеся в одном и том же микроинтервале времени после стимуляции при повторных предъявлениях движущегося стимула.
Рядом авторов показано, что при всем разнообразии признаков объекта, выделяемых зрительной системой различных животных для каждого отдельного вида существует свой видово-специфичный характер специализации зрительных нейронов, обеспечивающий главные для этого животного виды детекции. Для лягушки наиболее важными из зрительных детекторов являются, по-видимому, детекторы движения, а среди них - детекторы направления и скорости движения, позволяющие лягушке правильно соразмерять двигательную реакцию прыжка при ловле насекомых. Поэтому для исследования детекторных конструкций тектального уровня у лягушки целесообразно использовать нейроны, чувствительные к направлению и скорости движения маленького стимула, который в наших опытах имитировал движение темного жука по зеленому стеблю (темный “жук” - разрыв зеленой линии развертки, проходящей по диаметру экрана электронно-лучевой трубки; ее можно было переключать под углом в 45°, обеспечивая тем самым возможность движения “жука” в 8 направлениях и со скоростями 6÷60°/с) Исследование дирекциональной и скоростной чувствительности тектальных нейронов показало, что лишь немногие из них чувствительны только к направлению или скорости движения, а большинство реагирует на изменение обоих параметров движения, являясь таким образом, нейронами направления-скорости или “вектора скорости”. Возможно, такие нейроны являются интеграторами - “комплексными детекторами”, а возможно, они представляют собой проекции нейронов, первично чувствительных к обоим параметрам движения. Однако несмотря на то что преобладают нейроны направления-скорости (И. Кетлерис и др., 1973; I. Kettleris, A. Purtulite, 1974; и др., 1975, 1980; и др., 1976), для изучения характера переработки информации о движении целесообразно исследовать отдельно дирекциональную и скоростную чувствительности нейронов.
В наших опытах для выявления дирекциональной избирательности нейронов использовался стимул, движущийся в 8 направлениях (как было описано выше), со скоростью 18°/с, предъявляемых глазу 8-12-кратно с интервалами в 30 с. О дирекциональной избирательности судили по анализу векторных дифференциальных диаграмм. Для построения векторной диаграммы каждого нейрона использовалось 8 частотных гистограмм его импульсной активности, построенных соответственно движению стимула в 8 направлениях по которым затем строились две векторные диаграммы (одна - для фоновой активности, другая - для вызванной) на общей системе координат, представляющих собой лучи, расходящиеся из центра в 8 исследованных направлениях, на которых откладывались соответственно значения “плотности фона” и “плотности реакции”. Для определения характера избирательности нейрона к различным на правлениям движения объекта из каждой пары исходных векторных диаграмм строились разностные векторные диаграммы (без учета знака реакции), т. е. диаграммы, в которых координаты представляли модуль разности диаграмм вызванной и фоновой активности либо строились отдельно диаграммы В - и Т-реакций. Было обнаружено; что среди нейронов, чувствительных к направлению движения зрительного объекта, доминируют избирающие вертикальное направление (и особенно сверху вниз). По характеру и степени дирекциональной избирательности оказалось возможным выделить четыре типа нейронов: нейроны I типа, реагирующие на движение стимула только в одном из исследованных направлений (вернее, в секторе ±22,5°), их оказалось 12%; нейроны II типа, реагирующие на движение стимула в одном направлении значительно более ярко, чем в других, их оказалось 34%; нейроны III типа, реагирующие при движении объекта в двух и более (но не во всех) направлениях, их количество составило 36%; нейроны IV типа, не имеющие дирекциональной избирательности и реагирующие более или менее равнозначно при движении стимула во всех направлениях, их оказалось 18% от всех реагирующих на движение нейронов (рис. 3). Исследование распределения этих нейронов по слоям тектума лягушки, показало, что разной глубине их локализации соответствует преобладание определенного типа векторных диаграмм. Хотя и на большой и на малой глубинах встречаются нейроны всех типов, однако можно говорить о некотором преобладании нейронов, наиболее избирательных к направлению движения объекта (I, II типы) в более глубоких слоях (74% от всех нейронов на этой глубине, реагирующих на движение) и наименее дирекционально избирательных (III, IV типы) - в поверхностных (около 60%).
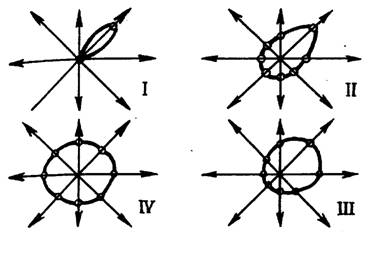
Рис. 3. Дифференциаторные диаграммы векторные диаграммы нейронов с разным типом дирекциональной чувствительности: I, II узкополосные; III, IV - широкополосные нейроны.
Наличие в тектуме лягушки нейронов с разным спектром дирекциональной избирательности, видимо, говорит о том, что выделение точного направления движения признака происходит с помощью процесса последовательного приближения, позволяющего вычислить наиболее вероятное направление движения зрительного Этот процесс происходит как в пределах каждого слоя, так и по пути из поверхностных слоев в глубокие, т. е. по ходу обработки зрительной информации. При этом если в поверхностных волокнистых и ядерных слоях, представляющих в основном волокна ГКС и в наружном ядерном и глубоком медуллярном слоях, представляющих собой основную массу сенсорных элементов, служащих проекций для ретинальных волокон, соотношение для разных типов нейронов сохраняется с преобладанием III и IV типов, обеспечивающих более общий зрительный анализ, то в глубоких внутреннем ядерном и внутреннем волокнистом, где наряду сенсорным анализом происходит интеграция и выдача команды на эффекторные нейроны, расположенные здесь же и в еще более глубоких структурах среднего мозга, соотношение между видами нейронов резко изменяется в сторону существенного преобладания I и II типов с обостренной дирекциональной избирательностью. Возможно также, что случаи, обнаруживающие векторные диаграммы I и II типов, свидетельствующие о тонком анализе в поверхностных слоях, связаны с наличием восходящих аксонных коллатералей из глубоких слоев в поверхностные (как и случаи ответов нейронов поверхностных слоев с длительными ЛП, описанные нами ранее).
Таким образом, можно полагать, что в КСМ лягушки в более поверхностно лежащих ретинальных элементах и сенсорных тектальных структурах, хотя и имеет место обострение зрительного анализа, способствующее послойному выделению направления движения, однако здесь более обширно представлены нейроны, способные к более общему зрительному анализу в отношении направления движения, констатирующие лишь наличие этого движения и в ряде случаев уточняющие направление его частотой импульсации, но не номером канала, в то время как в глубоких текталъных структурах происходит наибольшее обострение дирекциональной избирательности и направление движения кодируется уже номером канала, а внутри него может уточняться различными временными кодами. Такая интеграция, происходящая в глубоких тектальных слоях, позволяет, видимо, структурам, принимающим решение, группировать признаки, принадлежащие объекту наблюдения, и включать соответствующие эффекторные элементы (расположенные примерно на этой же глубине среди клеточных структур эфферентных слоев - , 1948) для осуществления биологической двигательной реакции нападения или избегания. Естественно, эта адекватная биологическая реакция определяется не только направлением движения стимула определенной величины и формы, но и в значительной степени скоростью его движения.
При анализе чувствительности тектальных нейронов к скорости движения стимула оказалось, что около 70% всех зарегистрированных нейронов обладает скоростной чувствительностью. По степени скоростной избирательности возможно выделить три группы нейронов: I группа - это нейроны, реагирующие на очень узкий диапазон скоростей (±3°), их можно рассматривать как узкополосные фильтры скоростей, количественно они составляют около 32% от всех реагирующих нейронов; II группа - это нейроны, реагирующие на более широкий диапазон скоростей (но не на все исследованные скорости) и описывающие отдельные скорости частотой импульсации и другими временными кодами, на долю этой группы приходится около 61% нейронов; III группу составили нейроны, реагирующие на весь диапазон исследованных скоростей и относящиеся также к описывающим нейронам, их оказалось около 7% (Т, В. Алейникова и др., 1977; , 1980; , , 1983). Наряду с различной степенью “скоростной избирательности” у тектальных нейронов отмечается чувствительность к различным участкам “скоростного спектра”, обеспечивающая различия в выборе конкретной скорости или определенного диапазона скоростей. Тектальные нейроны могут реагировать в довольно широких скоростных пределах, и при этом диапазоны их чувствительности могут перекрываться (рис. 4). Для тектальных нейронов, обладающих разными наборами скоростной чувствительности, наиболее оптимальными оказались скорости от 9 до 30°/с, а в данном диапазоне наибольшее количество реакций было зарегистрировано при движении зрительного стимула со скоростями 12 и 18° °/с.
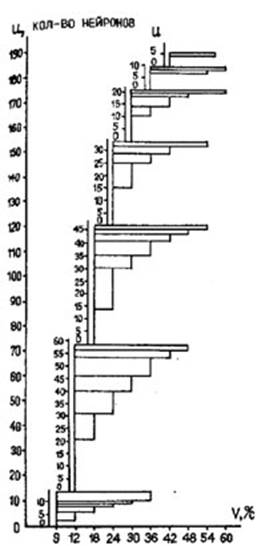
Рис. 4. Количественные соотношения исследованных тектальных нейронов, выделяющих различные скорости и диапазоны скоростей движения стимула: по оси абсцисс - предъявляемый набор скоростей движения стимула в °/с (v); по оси ординат - количество нейронов, реагирующих на данную скорость, и диапазон скоростей (u).
Несмотря на прослеживающуюся зависимость частоты импульсации нейронов от скорости движения зрительного стимула, при сравнении ответов нейронов с перекрывающимися диапазонами скоростной чувствительности видно, что одной частоты импульсации недостаточно для передачи информации о скорости движения. Большинство возбуждающихся нейронов КСМ лягушки, обладающих разными диапазонами скоростной чувствительности, кодируют скорость движения зрительного объекта в пределах диапазона своей чувствительности частотой импульсации, длительностью пачки импульсов, разностью ЛП ответов, повышая частоту, уменьшая длительность пачки, укорачивая ЛП и перестраивая паттерны ответа при увеличении скорости движения. И при этом диапазон анализируемых нейроном скоростей кодируется номером канала, а конкретная скорость внутри диапазона - частотно-латентно-числовым кодом ( и др., 1977; , 1980; и др., 1982; , , 1983). Для выявления “предпочитаемой” нейронами скорости был проведен корреляционный анализ зависимости реакций нейронов от скорости движения объекта наблюдения. Ответ нейрона на стимул нормировали по отношению к фоновой импульсации. На основании анализа связи между частотой импульсации нейрона и скоростью движения стимула были построены корреляционные уравнения, которые оказалось возможным разделить на 5 типов (рис. 5). К I типу были отнесены нейроны, экспериментальная зависимость между скоростью движения стимула и ответом может быть аппроксимирована отрезком прямой, параллельной оси абсцисс. Это нейроны чувствительные к изменению скорости движения зрительного объекта. При этом они либо вообще не реагируют на движение (аппроксимирующая прямая, параллельная оси абсцисс, проходит на уровне “I”, либо реагирует, но оказываются инваринтными к изменению скорости (в случае возбуждения аппроксимирующая прямая, параллельная горизонтальной оси, проходит на уровне выше “I”, а в случае торможения-ниже “I”). Таких нейронов I типа, безразличных к скорости движения, оказалось около 27%. Для нейронов II и III типов зависимость частоты импульсации от скорости движения также можно аппроксимировать прямыми, указывающими на линейную зависимость частоты импульсации в ответе нейрона от скорости движения зрительного стимула. Интересно, что эти нейроны не только могут увеличивать частоту импульсации (II тип) или уменьшать ее (III тип) при увеличении скорости, но и перестраивать свой ответ от тормозного до возбудительного (II тип) и, наоборот, от возбудительного до тормозного (III тип). По-видимому, нейроны II-III типов производят “вычисление” скорости движения, т. е. участвуют в описании скорости. Таких нейронов около 33%. У нейронов IV-V типов экспериментальная зависимость между скоростью движения стимула и ответом может быть аппроксимирована отрезком параболы. Нейроны этого типа отвечают на увеличение скорости движения сначала увеличением частоты (V тип), а затем уменьшением ее, либо наоборот, сначала уменьшением частоты (IV тип), а затем повышением ее. Нейроны IV-V типов предпочитают одну скорость из всего предъявляемого и анализируемого диапазона, отвечая максимальной В - (V тип) или Т - (IV тип) реакцией на движение объекта с определенной скоростью. Очевидно, нейроны IV-V типов можно считать детектирующими скорость движения зрительного объекта. Правда, анализ скорости приближается к детекторному типу не только при параболической зависимости, но еще и при условии, что ветви параболы поднимаются (или опускаются) круто. В противном случае, если ветви параболы изменяют свой наклон плавно, можно и эти нейроны отнести к описательным. Таких нейронов с параболической зависимостью частоты их импульсации от скорости движения зрительного стимула оказалось около 40%.
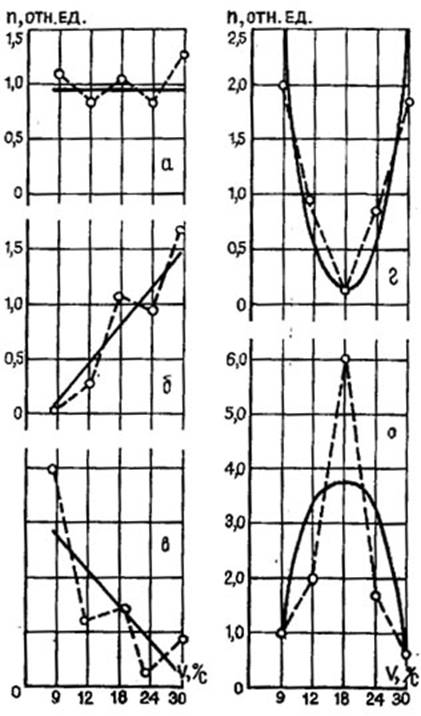
Рис. 5. Типы зависимостей частоты импульсной активности нейронов от скорости движения стимула: пунктирная линия - экспериментальная зависимость; сплошная - эмпирические модели; по оси абсцисс - скорости движения (v); по оси ординат - отношение числа импульсов в ответе к фоновому (n); а, б, в - I, II, III - типы зависимостей, которым удовлетворяет уравнение прямой; г, д, - IV, V - типы зависимостей, которым удовлетворяет уравнение параболы.
Таким образом, нейроны, чувствительные к скорости движения зрительного стимула, различаются между собой, во-первых, тем, что одни из них обладают широкой полосой пропускания скоростей, другие - относительно узкой, и, во-вторых, одни реагируют на изменение скорости повышением частоты импульсации в ответе, другие - снижением (нарастанием торможения). Естественно, возникает вопрос, не являются ли более узко настроенные нейроны более высоким уровнем зрительной системы и не формируется ли более узкая избирательность нейронов на базе нейронов, менее избирательных к скорости движения. В таком случае менее избирательных нейронов должно быть больше в том слое, куда приходит возбуждение из сетчатки, а более избирательных - в более глубоких слоях тектума.
В самом деле, в наших опытах отмечалась тенденция нарастания доли узкоизбирательных нейронов (I группа) в глубоких слоях тектума и уменьшение доли нейронов II группы. Распределение по глубине тектума нейронов с различной скоростной чувствительностью к разным участкам скоростного диапазона оказалось равномерным. Что касается нейронов с разными типами корреляционных уравнений, то здесь отмечается неравномерность распределения нейронов II и III типов по глубине тектума: количество нейронов этих типов достоверно уменьшается от поверхностных слоев к глубоким при повышении доли нейронов IV и V типов. Внутри каждого исследованного слоя количество нейронов разных типов также различно: в поверхностных слоях больше нейронов II и III типов, а в более глубоких - IV и V типов.
Таким образом, в тектальных структурах с их глубиной увеличивается удельный вес нейронов, более узкоизбирательных к направлению и скорости. При этом отмечается по мере нарастания глубины локализации нейронов достоверное увеличение количества Т - и уменьшение В-реакций, что, возможно, свидетельствует в пользу представления о наличии обострения избирательности к направлению и скорости движения в глубоких слоях КСМ. Факт наличия в тектуме лягушки нейронов с различными, но взаимно перекрывающимися диапазонами дирекциональной и скоростной чувствительности позволяет предположить непрерывность вычисления направления и скорости движения зрительного объекта в структурах КСМ. При этом детекция скорости обусловлена совместной деятельностью тектальных нейронов, функционирующих в совокупности как единая система измерения направления и скорости движения зрительного стимула. Количественное преобладание тектальных нейронов с широкими и перекрывающимися между собой диапазонами дирекциональной и скоростной чувствительности над нейронами, узкоспециализированными к детекции параметров движения, по-видимому, обеспечивает высокую надежность подобного процесса вычисления направления и скорости движения объекта.
То обстоятельство, что реакции нейронов II и III скоростных типов находятся в линейной зависимости от скорости движения объекта, но направлены реципрокно, может быть использовано в вычислении скоростей, как лежащих на границах предъявляемого диапазона, так и в средней его части, но механизм выделения скорости при этом должен быть разным. Так, детектирование скоростей, попадающих в средние, перекрывающиеся участки диапазонов, видимо, осуществляется с помощью механизма конвергенции импульсных потоков с нейронов II и III типов и пространственной суммации на нейронах-интеграторах, срабатывающих по определенному порогу. А выделение скоростей на границах чувствительности этих нейронов производится, вероятно, при малых скоростях за счет торможения активности нейронов II типа, обостряющего чувствительность нейронов III типа, а при больших скоростях, наоборот, торможение активности нейроов III тип способствует обострению В-реакций нейронов II типа. Реакции нейронов IV и V типов, имевшие также зеркальный характер, отражают нелинейную (параболическую) зависимость частоты импульсации нейронов от скорости движения стимула, и для каждого нейрона имеется некоторая оптимальная скорость движения, при которой он дает максимальную реакцию. При этом для разных нейронов эта оптимальная скорость была различной. В итоге были получены соответствующие корреляционные уравнения, охватывающие весь диапазон предъявляемых скоростей движения. Видимо, нейроны этих типов образуют совокупность единиц, остро чувствительных к определенным скоростям и участвующих, как и нейроны II и III типов, в классификации движущихся стимулов по скорости их движения. При анализе графиков корреляционных уравнений, характеризующих реакции II, III и IV, V типов, обращает на себя внимание тот факт, что графики корреляционных уравнений II и III типов могут быть участками ветвей параболы, характеризующей реакции нейронов IV и V типов, если для нейронов I и III типов предъявляемый нами диапазон скоростей был недостаточным для выявления всех возможностей этих нейронов; тогда при расширении диапазона скоростей характер кривой, возможно, мог измениться, если эти нейроны способны детектировать скорости, лежащие за пределами использованного нами диапазона. В пользу этого свидетельствуют некоторые “промежуточные” графики экспериментальных зависимостей импульсной активности от скорости движения, описываемые кривыми, приближающимися к параболе с несимметричными ветвями (рис. 6). По-видимому, определение истинной скорости движения зрительного стим6). Это может быть осуществлено отчасти, как и для определения направления, с помощью последовательного приближения к конкретному значению скорости за счет совокупной деятельности различных групп и типов нейронов с близкими и перекрывающимися диапазонами скоростной чувствительности.
Таким образом, на уровне tectum opticum лягушки, видимо, существуют два механизма вычисления наиболее вероятных направления и скорости движения: с помощью узкоизбирательных детекторов направления и скорости движения и с помощью широкополосных фильтров направлений и скоростей, допускающих аналоговое (не прерывное) описание любых адекватных для лягушки направлений и скоростей движения объекта.
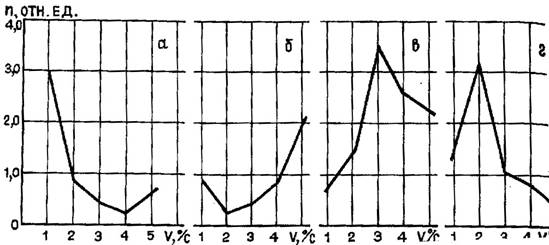
Рис. 6. Экспериментальные (“промежуточные”) типы зависимостей импульсной активности нейронов от скорости движения зрительного стимула: а, б - тип зависимости IV; в, г - тип зависимости V (кривые с несимметричными ветвями; обозначения те же, что и на рис. 5)
Такое исследование скоростной либо дирекциональной чувствительности нейронов, проводимое раздельно, позволяет изучить данные их свойства, но искусственно их разрывает в тех случаях, когда они оказываются тесно связанными. Так, исследования последних лет показали, что кроме нейронов, чувствительных к направлению движения и к изменению скорости движения есть вид нейронов, которые реагируют изменением импульсной активности при изменении и скорости, и направления движения. Это нейроны, описанные И. Кетлерисом и А. Пуртулите (1974) для коры кролика как нейроны скорости-направления и нами ( и др., 1976; и др., 1979) как детекторы вектора скорости или как нейроны направления скорости. Такие нейроны, будучи чувствительны к нескольким (по крайней мере к двум) параметрам движения, видимо, участвуют в интегральном описании образа и, вероятно, обусловливают в основном характер ответной двигательной реакции животного. Для исследования таких комплексных дирекционально-скоростных свойств нейронов использовалось перемещение стимула в разных направлениях и с разными скоростями, в результате чего была показана ведущая роль нейронов, выделяющих направление и скорость, в “вычислении" параметров движения зрительного объекта, о чем свидетельствует их большой удельный вес (91% от числа нейронов, реагирующих на движение) по сравнению с только “дирекциональными” нейронами (около 5%) и только “скоростными” (около 4%), о возможной роли которых следует, видимо, говорить с осторожностью из-за малого их. количества.
Таким образом, показано, что более половины тектальных нейронов участвует в детекции признаков формы, размера, направления и скорости движения стимула. Некоторые из этих нейронов работают как узкополосные фильтры, остро выделяя только один признак, но большинство функционирует подобно широкополосным фильтрам, описывая образ различными импульсными кодами, т. е. для “вычисления” наиболее вероятных направления, скорости движения, размера объекта используются две системы - дискретная детекторная и непрерывная - описательная. Все это, естественно, повышает надежность передачи информации.
1.3. РЕЗЮМЕ
В главе рассматриваются некоторые функционально-морфологические характеристики нейронов ретино-тектальной системы лягушки, реагирующих на зрительную стимуляцию, в том числе конструкция их рецептивных полей и детекторные функции. В результате исследования, во-первых, показано, что большинство ретинальных нейронов имеют стабильные латентные периоды ответов на стереотипную стимуляцию, изменяющиеся в зависимости от интенсивности воздействия; некоторые ГКС имеют константный ЛП, сохраняющийся при изменении характера стимуляции, что позволяет считать эти нейроны “синхронизаторами”, обеспечивающими наличие “точки отсчета” при использовании в качестве кандидата в коды разности ЛП ответов нейронов. Во-вторых, оказалось, что разность ЛП реакций может быть информативной и в работе тектальных нейронов, перерабатывающих и передающих зрительные сигналы. В-третьих, выяснилось, что большинство зрительных тектальных нейронов являются поливалентными, реагируя (хотя и по-разному) на различные формы зрительных воздействий. В-четвертых, отмечается существенная роль торможения в ответах тектальных нейронов, способствующего, видимо, обострению возбудительной реакции анализирующих воздействие нейронов тектума. В-пятых, обсуждаются вопросы организации рецептивных полей тектальных нейронов; показано, что в тектуме преобладают гетерогенные поля, часто с перекрывающимися возбудительными и тормозными зонами, круглые и вытянутые, среди которых особое место занимают РП с характерной комбинацией мелких В - и Т-зон, образующих сложный мозаичный узор поля; это, видимо, позволяет таким “мозаичным” полям выполнять роль анализаторов движущихся мелких зрительных стимулов-объектов. В-шестых, отмечается значительная вариабельность характера реакций ряда тектальных нейронов и ЛП их ответов при повторной стимуляции вплоть до прекращения реагирования (нейроны “новизны”) или до появления опережающих стимул реакций (нейроны “тождества” и среди них “экстраполяционные нейроны”). В-седьмых, показано, что движение возбуждения по тектальным нейронам (“цепочки возбуждения”) в случае действия разных стимулов имеет различный характер. В-восьмых, обнаружено, что более половины нейронов тектума обладает детекторными функциями, которые связаны, как и в сетчатке, с выделением контраста, выпуклых и движущихся краев, затемнения, размеров объекта, направления и скорости его движения; при этом на уровне тектума по сравнению с сетчаткой увеличивается количество нейронов, дирекционально избирательных и селективных к скорости движения зрительного объекта. В-девятых, показано равномерное распределение по слоям тектума нейронов, связанных с различными детекторными функциями, но при этом отмечается обострение дирекциональной избирательности и скорости селективности в глубоких слоях КСМ, где формируется команда к действию по сравнению с поверхностными, отражающими ретинальную неизбирательность к направлению и скорости движения; обнаружено, что основная группа нейронов, анализирующая параметры движения, представляет собой нейроны направления-скорости, реагирующие наилучшим образом при движении стимула в оптимально направлении (для данного РП) с оптимальной скоростью. В-десятых, по характеру и степени селективности к направлению и скорости нейроны образуют несколько групп, причем самые избирательные подобны узкополосным фильтрам и являются детекторами, вычленяющими “ключевые” признаки сигнала, а менее избирательные действуют как широкополосные фильтры, работающие по принципу описательных систем; те и другие элементы образуют надежную систему “вычисления” наиболее вероятных направления и скорости движения зрительного стимула.
ГЛАВА 2
КОДИРОВАНИЕ И ПЕРЕРАБОТКА ЗРИТЕЛЬНОЙ ИНФОРМАЦИИ В НЕТЕКТАЛЬНЫХ ОПТИЧЕСКИХ СТРУКТУРАХ МОЗГА ЛЯГУШКИ
У амфибий кроме основной тектальной имеются еще две оптические системы: добавочная ретино-тегментальная, образованная базальным оптическим трактом и ядром базального оптической тракта (, 1968; W. Riss et al., 1963), и ретино-таламо-теленцефальная, которая в результате эволюционного развития мозга становится ведущей, а у лягушки обеспечивает опознание стационарных стимулов.
2.1. МЕЗЕНЦЕФАЛЬНЫИ УРОВЕНЬ. ТЕГМЕНТУМ
Покрышка (tegmentum) ножки среднего мозга бесхвостых амфибий содержит ряд ядер и путей, основными из которых, по (1968), являются следующие: ядро глазодвигательного нерва, моторное ретикулярное ядро и ядро базального оптического тракта. Большинство ядерных образований покрышки относится к ретикулярной формации среднего мозга. По данным ряда авторов (W. Riss et al., 1963; , 1968; , 1974; , 1977), тегментум имеет многочисленные связи с разными отделами мозга. Так, описаны связи покрышки среднего мозга с его крышей (, 1968) и передним мозгом (, 1977), с продолговатым и спинным мозгом (, 1968). В тегментум поступают импульсы различной сенсорной модальности: обонятельные, акустические, соматические, зрительные (, 1974). Есть данные о том, что в покрышке среднего мозга низших позвоночных, в частности амфибий, оканчиваются прямые зрительные афференты от сетчатки, которые образуют один из трех пучков зрительного тракта: базальный оптический тракт, относящийся к добавочной оптической системе (, 1968; , 1972; , 1974; , 1976; , 1977; , 1980; , , 1982). Полагают, что роль добавочной оптической системы - в передаче сигналов зрительной модальности на моторные ядра тегментума для осуществления двигательных реакций (W. Riss et al., 1963; , 1968; , 1974; , 1976). Базальный оптический тракт (БОТ) отходит от базальной части хиазмы и направляется дорсокаудально. Латеральная часть волокон БОТ входит в ядро БОТ (ЯБОТ) с вентральной стороны. Медиальная часть БОТ входит в ЯБОТ с передней и вентральной стороны. В составе БОТ насчитывается 600-800 миелинизированных и не-миелинизированных волокон. Все миелинизированные волокна являются аксонами ГКС (, 1977).
В исследованиях на травяной лягушке (, 1968) описано ЯБОТ, локализованное в вентральной части покрышки среднего мозга и имеющее овальную форму, немного вытянутую в рострокаудальном направлении. Клетки ЯБОТ различаются по величине в зависимости от локализации в ядре: в дорсальных отделах ядра располагаются клетки средних размеров, в вентральных и медиальных - более мелкие. По форме они практически не отличаются, имеют крупные ядра с небольшим количеством хроматина, окруженные узким ободком цитоплазмы. Вентральная часть покрышки богата кровеносными сосудами, что косвенно свидетельствует о большой Физиологической значимости этой области мозга. С помощью односторонней перерезки зрительного нерва по перерождению зрительных

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
