

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
МИНИСТЕРСТВО ВНУТРЕННИХ ДЕЛ РОССИЙСКОЙ ФЕДЕРАЦИИ КРАСНОДАРСКИЙ УНИВЕРСИТЕТ МВД РОССИИ
КАФЕДРА ПСИХОЛОГИИ И ПЕДАГОГИКИ
«У Т В Е Р Ж Д А Ю»
Начальник кафедры
подполковник полиции
______________
«___» ___________ 2012 г.
Дисциплина: «Анатомия и физиология центральной нервной системы ».
По направлению подготовки (специальности) 050407.65 Педагогика и психология девиантного поведения
специализация - Социальная педагогика
ЛЕКЦИЯ
ТЕМА 6: «Внешнее и внутреннее строение продолговатого, заднего, среднего мозга»
Обсуждена и одобрена на заседании кафедры Протокол № от «___»_________20 г. | Подготовил: Начальник кафедры психологии и педагогики
|
Краснодар 2012
Объем времени, отводимого для изучения данной темы: 4 часа.
Место проведения: учебная аудитория.
Методика проведения: классическая лекция.
Основное содержание темы:
Образование заднего мозга. Строение заднего мозга, его элементы (варолиев мост, а также мозжечок) и их топография. Поперечные пучки сформированы отростками нейронов, входящими в ядра моста. Они направляются в сторону мозжечка, образуя средние мозжечковые ножки, задней части моста кроме волокон восходящего направления, которые являются продолжением чувствительных проводящих путей продолговатого мозга. Находят очаговые скопления серого вещества, а именно ядра V, VI, VII, VIII пар.
Основные термины и понятия:
Кора головного мозга, мозжечок, варолиев мост, средние мозжечковые ножки, серое вещество, узлы (ядра) в толще белого вещества, базальные ядра или подкорковые центры, полосатое тело, ограда, миндалевидное тело.
Цели занятия:
1. Формирование у курсантов, слушателей прочных знаний о строении базальных ядер.
2. Закрепить знания в области строения таламической области.
3. Содействовать осознанию курсантами, слушателями важности изучаемого предмета.
Вопросы занятия:
1.Строение мозжечка и моста.
2.Строение продолговатого мозга.
3 Строение среднего мозга
Литература
1. , , Фельдман центральной нервной системы. – Ростов-на-Дону: Феникс, 2006.
2. , Цехмистренко нервной системы. – М.: Мир, 2006.
3. Физиология человека: учебник для мед. ин-тов / под ред. . М.: 2009.
4. , Туровский центральной нервной системы для психологов. – СПб.: Питер, 2009.
5. , Кульба центральной нервной системы. Р-н-Д 2008 г.
Дополнительная литература
1. Антонова и физиология центральной нервной системы. – М.: Высшее образование, 2006.
2. , Туровский центральной нервной системы для психологов - СПб.: Питер, 2007
3. Дерягина физиологии центральной нервной системы. – М.: Изд-во МосУ МВД России, 2008.
4. Анатомия человека (под ред. Сапина). М., 1986, Т.2
5. , и др. Физиология ЦНС. - Ростов-на-Дону: Феникс, 2000
6. Физиология: Основы и функциональные системы: Курс лекций / Под. ред. Судакова.- М., 2000.
7. , Атлас анатомии человека. 1992, Т. 4.
1. Продолговатый мозг
ВНЕШНЯЯ ФОРМА
Продолговатый мозг, является непосредственным продолжением спинного мозга. Он имеет форму усеченного конуса, обращенного основанием вверх. Его средняя длина — 25 мм, наибольшая ширина у основания — 22 мм, толщина — 14 мм, средняя масса — около 6 г. Своей вентральной поверхностью продолговатый мозг прилегает к нижней части ската затылочной кости и к зубу II шейного позвонка. Чтобы понять строение продолговатого мозга, необходимо вспомнить, что по развитию в онтогенезе он представляет собой сегмент нервной трубки. Однако в процессе органогенеза боковые стенки первоначальной нервной трубки становятся значительно толще, а дорсальная стенка истончается и сохраняется лишь в виде тонкой пластинки. Она представляет собой эпендимальный эпителий, к которому прилегает снаружи сосудистая оболочка IV желудочка.
Продолговатый мозг имеет вентральную, дорсальную и боковые поверхности.
На вентральной поверхности продолговатого мозга располагается передняя срединная щель, которая является непосредственным продолжением такой же щели спинного мозга.
По обе стороны от щели располагаются два продольных валика. Это пирамиды, образованные нервными волокнами нисходящего направления. На границе продолговатого мозга со спинным мозгом большая часть волокон каждой пирамиды переходит на противоположную сторону, в результате образуется перекрест пирамид (продолг. вентр). По перекресту пирамид определяется нижняя граница продолговатого мозга. Латеральнее пирамид находится передняя латеральная борозда, которая является продолжением одноименной борозды спинного мозга. Эта борозда хорошо различима в верхней части органа, где из нее выходят корешки подъязычного нерва — XII пара. В нижней части борозда прерывается поперечно идущими наружными дугообразными волокнами.
Из верхней части передней латеральной борозды выступает овальной формы возвышение, называемое оливой. Олива, имеет длину около 14 мм и ширину 5 мм. Ее верхний конец находится вблизи моста. Латеральнее оливы находится задняя латеральная борозда продолговатого мозга, которая не соответствует задней латеральной борозде спинного мозга. Из этой борозды на продолговатом мозге выходят краниальные корешки добавочного нерва — XI пара, корешки блуждающего нерва — X пара и языкоглоточного нерва — IX пара.
Дорсальная поверхность продолговатого мозга имеет различное строение в нижней и верхней частях (продолг. дорсальн).
По сторонам от срединной борозды в нижней части располагаются правый и левый задние канатики. Примерно на середине длины продолговатого мозга они расходятся латерально и вверх, а затем продолжаются в виде толстых валиков, называемых нижними ножками мозжечка, которые погружаются в мозжечок. Между правой и левой нижними ножками мозжечка образуется площадка треугольной формы, которая представляет собой нижнюю половину ромбовидной ямки.(ромб. ямка). В месте отхождения задних канатиков от срединной линии составляющие их тонкий и клиновидный пучки имеют небольшие утолщения, называемые соответственно тонким и клиновидным бугорками.
ВНУТРЕННЕЕ СТРОЕНИЕ
(прод мозг) В образовании продолговатого мозга принимают участие как серое, так и белое вещество.
Серое вещество, в отличие от спинного мозга, теряет форму бабочки, распределяется неравномерно и представлено четырьмя группами ядер.
Волокна корково-спиномозгового пути занимают место в переднем канатике спинного мозга на своей стороне, образуя передний корково-спинномозговой тракт.
Вблизи дорсальной поверхности продолговатого мозга по сторонам от срединной плоскости находится задний продольный пучок, вентральнее от него — медиальный продольный пучок. Кпереди от последнего располагается покрышечно-спинномозговой путь.
Медиальнее афферентного спинно-таламического пути расположен красноядерно-спинномозговой путь. Дорсальнее пирамид проходит ретикулярно-спинномозговой путь и латеральнее от него — пред-дверно-спинномозговой путь.
Помимо проходящих транзитом через продолговатый мозг афферентных и эфферентных путей имеются тракты, которые связывают чувствительные ядра IX и X пар черепных нервов с интеграционными центрами головного мозга — ядерно-таламический путь и ядерно-мозжечковый путь.
Ядерно-таламический путь является трактом общей чувствительности (поверхностной и глубокой) от области головы. Кроме того, он обеспечивает доставку информации от интероцепторов (рецепторов внутренних органов). Ядерно-мозжечковый путь проводит бессознательные проприоцептивные импульсы от области головы. На нейронах двигательных ядер IX, X, XI и XII пар черепных нервов заканчиваются волокна корково-ядерного пути.
Учитывая гистотопографические отношения в продолговатом мозге, следует отметить, что в пирамидах проходят нисходящие пирамидные тракты. Они начинаются от эффекторных клеток коры полушарий большого мозга, осуществляют сознательные (по желанию человека) движения и оказывают тормозное воздействие на сегментарный аппарат ствола головного и спинного мозга.
В покрышке продолговатого мозга находится серое вещество — ядра указанных черепных нервов, ретикулярная формация, ядра оливы и белое вещество — проводящие пути.
В ретикулярной формации продолговатого мозга располагаются жизненно важные центры: дыхательный; сосудодвигательный; центры слюноотделения, глотания, секреции желудочного, панкреатического, кишечного соков и центры защитных рефлексов (рвота, кашель, чихание). Ядра оливы отвечают за безусловно-рефлекторную регуляцию тонуса мышц при вестибулярных нагрузках. Кроме того, внутри продолговатого мозга проходят афферентные (восходящие) и эфферентные (нисходящие) экстрапирамидные тракты. Последние начинаются от подкорковых двигательных центров и отвечают за безусловно-рефлекторные движения и тонус мышц.
Дорсальная поверхность продолговатого мозга образует нижний отдел дна четвертого желудочка (ромбовидной ямки).
Волокна X и XII пар черепных нервов разделяют поперечный срез продолговатого мозга на три отдела с каждой стороны — медиальный, латеральный и дорсальный. Медиальный отдел ограничивает передняя срединная щель и волокна подъязычного нерва. Латеральный отдел располагается между волокнами подъя-зычного и блуждающего нервов. Дорсальный отдел находится кнару-жи от волокон блуждающего нерва.
Знание локализации указанных структур важно для понимания функциональных нарушений, наблюдающихся в неврологической практике при различных повреждениях.
Расстройства функций продолговатого мозга именуют бульбар-ными, исходя из синонима: продолговатый мозг — луковица мозга. Они характеризуются нарушениями дыхания, сердечной деятельности, тонуса сосудов, нарушениями глотания и функций черепных нервов XII—IX пар. Эти расстройства могут привести к смерти.
Ретикулярная формация
В толще продолговатого мозга расположена группа нейронов со сложно переплетающимися волокнами. Подобные нейроны обнаруживаются также в верхнешейных сегментах спинного мозга, в среднем и даже промежуточном мозге. Под микроскопом подобное образование напоминает сеть. Отсюда и название — ретикулярная формация. Можно выделить «белую» ретикулярную формацию (с преобладанием миелинизированных волокон) и «серую» ретикулярную формацию (состоящую из клеток и слабо миелинизированных волокон).
Ретикулярная формация образована группами мелких, средних и крупных мультиполярных вставочных нейронов с различным характером ветвления дендритов и аксонов, содержащих различные нейромедиаторы и окруженных сетями нервных волокон. При этом диффузное расположение ее элементов сменяется более компактным с формированием отдельных ядер, которых насчитывается большое количество.
Нейроны ретикулярной формации характеризуются большим количеством афферентных связей, идущих от сенсорных образований, и богатством эфферентных проекций. Их отростки направляются в кору больших полушарий, в ядра различных отделов ствола, конечного мозга и мозжечка, объединяя их в одну систему (в этом проявляется интпегративная функция ретикулярной формации). Обширные восходящие проекции, в том числе непосредственно в кору большого мозга, обеспечивают активирующее влияние ретикулярной формации на высшие центры нервной системы. Совокупность нисходящих проекций ретикулярной формации рассматривают как систему, угнетающую активность нижележащих центров. Основной объем этих проекций представлен волокнами ретикулоспинального тракта, которые угнетают активность мотонейронов спинного мозга. Ретикулярной формации приписывают участие в восприятии боли, агрессивном и половом поведении. Основными медиаторами в ней являются ацетилхолин, норадреналин, дофамин, серотонин.
Ретикулярная формация получает сенсорные сигналы по различным путям и действует как своего рода фильтр, пропуская только ту информацию, которая является новой или необычной. Волокна от нейронов ретикулярной системы идут в различные области коры больших полушарий, некоторые — через таламус. Считается, что большинство нейронов ретикулярной формации являются «неспецифическими». Это означает, что, в отличие от нейронов первичных сенсорных путей, например зрительных или слуховых, реагирующих только на один вид раздражителей, нейроны ретикулярной формации могут реагировать на многие виды стимулов. Эти нейроны передают сигналы от глаз, кожи, внутренних органов, а также других органов и структур лимбической системе к коре.
Исследователи выяснили, что некоторые участки ретикулярной формации обладают более определенными функциями. Таково, например, голубое пятно — плотное скопление тел нейронов, отростки которых образуют дивергентные сети с одним входом, использующие в качестве медиатора норадреналин. Часть нервных путей идет вверх от голубого пятна к таламусу, гипоталамусу, многим областям коры. Другие направляются вниз, к мозжечку и спинному мозгу. Норадреналин — медиатор этих специализированных нейронов — может запускать эмоциональные реакции. Было высказано предположение, что недостаток норадреналина в мозгу приводит к депрессии, а при длительном избыточном воздействии норадреналина возникают тяжелые стрессовые состояния. Возможно, норадреналин играет также роль в возникновении реакций, субъективно воспринимаемых как удовольствие. Другой участок ретикулярной формации — «черная субстанция» — представляет собой скопление тел нейронов, опять-таки принадлежащих к дивергентным сетям с одним входом, но выделяющих медиатор дофамин.
(строение ГМ) Исходя из онтогенетической классификации отделов головного мозга, задний мозг включает в себя: мост, мозжечок, перешеек ромбовидного мозга, продолговатый мозг.
Задний и продолговатый мозг образовались в результате деления ромбовидного мозгового пузыря. Полостью заднего мозга является IV желудочек.
МОСТ
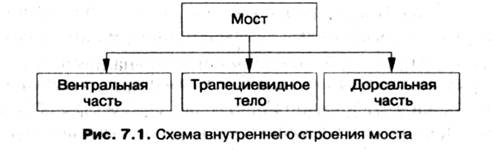
(мост с вентр. стор) Мост расположен выше продолговатого мозга и является его продолжением. Спереди мост имеет вид поперечно расположенного валика, граничащего сверху со средним мозгом, а снизу — с продолговатым мозгом. Его длина по срединной линии составляет 2,5 см, ширина 3—3,5 см, толщина 2,5 см, масса — 16—18 г. Латерально мост продолжается в средние ножки мозжечка. Дорсальная поверхность моста участвует в образовании дна IV желудочка, ему принадлежит верхний угол ромбовидной ямки. На поперечном срезе моста выделяют три слоя: спереди — базилярную часть, сзади — покрышку и разделяющее их трапециевидное тело.
В покрышке (дорсальная часть) проходят афферентные проводящие пути, а также залегают клетки ретикулярной формации. В ретикулярной формации моста так же, как и в продолговатом мозге, расположены жизненно важные центры, в том числе центры дыхания.
Трапециевидное тело представляет собой пучок поперечно идущих нервных волокон, между которыми имеются клеточные скопления ядра трапециевидного тела. Большая часть волокон берет начало в ядрах преддверно-улиткового нерва и переходит на противоположную сторону, формируя перекрест.
Базилярная часть (вентральная часть) является филогенетически молодым образованием моста, она появилась для соединения коры головного мозга через средние ножки с мозжечком. В ядрах моста нервные импульсы из коры переключаются на нейроны, чьи аксоны идут к мозжечку через средние мозжечковые ножки (рис. 7.1).
(ГМ с вентр. стор) В поперечной борозде, отделяющей мост от пирамид продолговатого мозга, выходят корешки правого и левого отводящих нервов (VI пара черепно-мозговых нервов) боковой части этой борозды видны корешки лицевого нерва (VII пара). На передней поверхности моста заметна широкая, но не глубокая борозда с одноименной артерией. Внутреннее вещество моста неоднородно. В центре заметен толстый пучок волокон расположенный поперечно и относящийся к проводящим путям слухового анализатора (трапециевидное тело). Это образование делит мост на заднюю и переднюю часть (покрышечную и базилярную).
Между волокнами трапециевидного тела расположены ядра передние и задние. В передней части моста видны продольные и поперечные волокна. Продольные пучки моста относятся к пирамидному пути и волокнам, соединяющим корковые спинномозговые центры, образующие отдельны пучки. В этой же группе волокон располагаются корково-мозговые волокна, которые начинаются в корковых центрах и заканчиваются в ядрах моста. Они расположены группами в толще моста. Поперечные пучки сформированы отростками нейронов, входящими в ядра моста. Они направляются в сторону мозжечка, образуя средние мозжечковые ножки, задней части моста кроме волокон восходящего направления, которые являются продолжением чувствительных проводящих путей продолговатого мозга.
Основание моста образовано серым и белым веществом. Серое вещество основания моста представлено многочисленными мелкими собственными ядрами моста. К нейронам этих ядер подходят и на них заканчиваются синапсами нервные волокна, происходящие от нейронов коры полушарий большого мозга — корково-мостовой путь. Аксоны нейронов этих ядер направляются на противоположную сторону и в составе средних ножек мозжечка достигают коры полушарий мозжечка. Этот участок проводящего пути носит название мосто-мозжечковый тракт.
Белое вещество основания моста представлено волокнами, имеющими продольное и поперечное направления. Транзитом проходит продольно ориентированный корково-спинномозговой тракт, который в пределах моста представлен разрозненными пучками волокон. Только в области нижнего края моста они собираются в компактный пучок и затем образуют пирамиды продолговатого мозга. Такое же продольное направление в основании моста имеют волокна корково-ядерного пути, который частично заканчивается на нейронах двигательных ядер V, VI и VII пар черепных нервов, а затем продолжается к двигательным ядрам IX—XII пар черепных нервов, расположенных в продолговатом мозге.
Наконец, продольное направление имеют уже упомянутые волокна tr. corticopontinus, заканчивающиеся на нейронах собственных ядер моста. Аксоны нейронов названных ядер обусловливают поперечную исчерченность вентральной поверхности моста и составляют мосто-мозжечковый путь.
Дорсальная часть моста — его покрышка, также содержит серое и белое вещество. Серое вещество покрышки представлено ядрами V, VI, VII и VIII пар черепных нервов, которые находятся в основном в дорсолатеральной части покрышки. Кроме того, здесь имеются ядра, входящие в состав ретикулярной формации, расположенной по обе стороны от срединной плоскости.
Тройничный нерв, V пара черепных нервов, имеет ядра не только в пределах моста. Они располагаются также в среднем мозге и в спинном мозге. В покрышке моста находятся чувствительное мостовое ядро, или верхнее чувствительное ядро, и двигательное ядро тройничного нерва. Мостовое ядро тройничного нерва является коммуникационным центром проводящего пути тактильной чувствительности от области лица.
МОЗЖЕЧОК
Мозжечок, cerebellum, развивается из дорсальной стенки заднего мозга и является самой крупной, после полушарий большого мозга, частью головного мозга. В функциональном отношении мозжечок является интеграционным центром ромбовидного мозга — центром статокинетических и вестибулярных функций.
Вместе с продолговатым мозгом и мостом мозжечок располагается в задней черепной ямке.
Мозжечок имеет ромбовидную форму с преобладанием поперечного размера. Его ширина в среднем равняется 10 см, длина по срединной линии 3—4 см, толщина 4—5 см; средняя масса около 135 г.
В мозжечке выделяют среднюю часть — червь, две боковые объемистые части — полушария. Исходя из развития мозжечка в филогенезе, следует выделить небольшое образование, прилегающее с вентральной стороны к полушарию — клочок. В черве и полушариях мозжечка различают две поверхности: верхнюю и нижнюю.
Верхняя поверхность мозжечка, обращена вверх и назад. Она выпуклая и посередине имеет продольное возвышение, называемое верхним червем. Червь с латеральных сторон переходит в полушария. Нижняя поверхность мозжечка, направлена вниз и вперед. Она прилежит к затылочной кости. На нижней поверхности мозжечка имеется продольное углубление, называемое долинкой. В этом углублении находится нижний червь, отделенный от каждого полушария бороздой.
Поверхность мозжечка исчерчена большим количеством параллельных друг другу борозд, которые имеют поперечное направление и различную глубину. Мелкие борозды разделяют поверхность мозжечка на пластиночки, более глубокие борозды разделяют группы пластиночек на пластины, которые получили название листки мозжечка. Наконец, самые глубокие борозды разделяют поверхность мозжечка на дольки.
Среди борозд, разделяющих дольки мозжечка, самой глубокой (до 2 см) является горизонтальная борозда. Она проходит по всей окружности мозжечка и разделяет верхнюю и нижнюю поверхности полушарий.
Борозды мозжечка, не прерываясь, переходят от червя на полушария. Клочок также имеет пластинки, но дольки в нем не выделяют.
В черве различают восемь долек, по четыре дольки в верхнем и нижнем черве. Самой передней долькой верхнего червя является язычок. Эта долька образована несколькими пластинками.
Такое разделение мозжечка на дольки было дано на основе предпосылки наличия связей между отдельными частями полушарий и определенными участками червя. Современные исследования проводящих путей мозжечка позволяют считать более рациональным выделение частей, функция которых формировалась в процессе фило - и онтогенеза. Так, в мозжечке выделяют филогенетически древнюю часть (paleocerebellum), которая включает в себя клочок и узелок, старую часть (archicerebellum), к которой относятся червь, за исключением узелка, и новый мозжечок (neocerebellum), включающий в себя полушария мозжечка, развивающиеся из средней части червя.
Посредством трех пар ножек мозжечок соединен с различными отделами головного мозга: верхними — со средним мозгом, средними—с мостом, нижними — с продолговатым мозгом.
ВНУТРЕННЕЕ СТРОЕНИЕ
На разрезах можно видеть расположенное на поверхности серое вещество, образующее кору мозжечка, под корой мозжечка — белое вещество, составляющее мозговое тело, от которого к поверхности тянутся отростки, проникающие в дольки и пластинки мозжечка. На срединном разрезе мозжечка белое вещество имеет листовидную форму, с которой связано образное название «древо жизни».
В толще белого вещества мозжечка располагаются скопления серого вещества, составляющие ядра мозжечка. В черве мозжечка по обе стороны от срединной линии находится ядро шатра. Латеральнее этого ядра находится второе небольшое ядро, называемое шаровидным. Еще более латерально лежит пробковидное ядро. В белом веществе полушарий находится самое крупное ядро – зубчатое. Ядро шатра относится к древнему мозжечку, шаровидное и пробковидное ядра являются филогенетически более поздними образованиями и относятся к старому мозжечку, а зубчатое ядро относится к новому мозжечку.
Белое вещество мозжечка содержит афферентные и эфферентные волокна, которые связывают мозжечок со стволом мозга и формируют ножки мозжечка. Различают три пары ножек мозжечка: верхние, средние и нижние. Верхние ножки мозжечка связывают его со средним мозгом, средние ножки - с мостом, нижние ножки – с продолговатым мозгом.
ПЕРЕШЕЕК
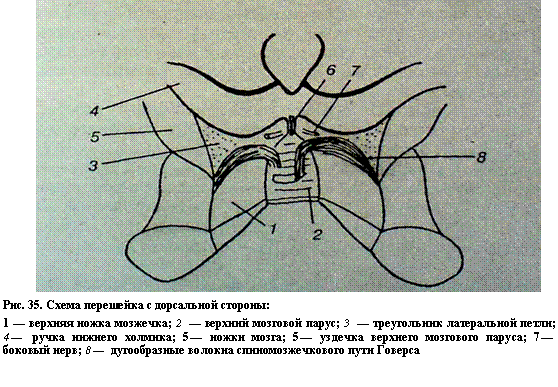
Перешеек ромбовидного мозга представляет собой небольшой участок, лежащий между ромбовидным и средним мозгом. Из этой части развиваются верхние ножки мозжечка, верхний мозговой парус, треугольник слуховой петли. Верхние ножки мозжечка начинаются от его ядер и заканчиваются у края нижнего двухолмия среднего мозга. Между ножками натянут верхний мозговой парус состоящий из эпендимальных клеток. Впереди и латерально от конца верхних ножек мозжечка на боковой поверхности ножек мозга имеется треугольная площадка. В этом треугольнике пучок волокон слуховой петли приближается к поверхности ромбовидного мозга (рис. 35).
Введение
Средний мозг (mesencephalon) развивается в процессе филогенеза под преимущественным влиянием зрительного рецептора, поэтому важнейшие его образования имеют отношение к иннервации глаза. Здесь же образовались центры слуха, которые вместе с центрами зрения в дальнейшем разрослись в виде четырех бугорков — четверохолмие. С появлением у высших животных и человека слухового и зрительного анализаторов в коре переднего мозга слуховые и зрительные центры среднего мозга попали в подчиненное положение и стали промежуточными, подкорковыми. С развитием у высших млекопитающих и человека переднего мозга через средний мозг стали проходить проводящие пути, связывающие кору концевого мозга со спинным (ножки мозга).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
