

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
С. Н. Боранбаев, Д. А. Саранча, Р. Таберхан,
Применение комбинированного метода для создания математических моделей биогеоценозов различных регионов казахстана (имитационная модель «растительность - лемминги - песцы»)

(Евразийский национальный университет имени , г. Астана, (Вычислительный Центр Российской Академии Наук, г. Москва)
Рассматривается имитационная модель «растительность - лемминги - песцы». Решается «обратная имитационная задача». Приведен список основных параметров для моделирования животных (в том числе хищников).
Описание имитационной модели «растительность - лемминги - песцы»
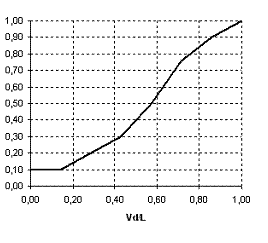
Модель описывает динамику численности песцов (F), леммингов (L), и их корма (V). Влияние времени года на динамику описывается с помощью вспомогательных функций. Зимой:  Весной
Весной ![]()
![]() . Летом:
. Летом: .
.
Таблица 1.
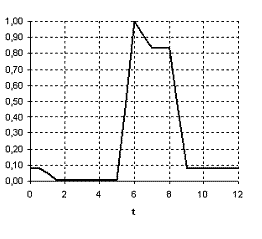
Время года | зима | весна | лето | зима | ||||||||
месяц | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
S1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
S2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
|
|
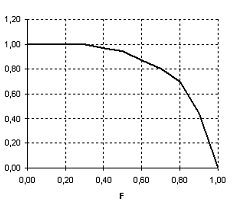
Рисунок 1. Функция, формализующая снижение ценности кормов при их дефиците (g2) | Рисунок 2. Функция зависимости смертности леммингов от сезона (g3) |
|
|
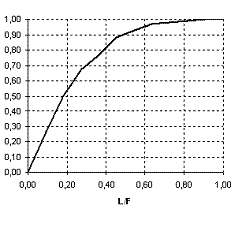
Рисунок 3. Функция насыщения выедания леммингов песцами (g4) | Рисунок 4. Функция самолимитирования песцов (g5) |
В течение зимы и весны часть растительности ![]() находится под снегом, и поэтому перестает взаимодействовать с леммингами и меняется только в соответствии со своей собственной динамикой. С наступлением лета части растительности над и под снегом объединяются.
находится под снегом, и поэтому перестает взаимодействовать с леммингами и меняется только в соответствии со своей собственной динамикой. С наступлением лета части растительности над и под снегом объединяются.
| (1) |
| (2) |
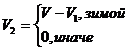 ( растительность под снегом)
( растительность под снегом)Но лемминги выедают только часть растительности, в основном надземную, которая составляет долю ![]() от ее общей биомассы. Если биомасса доступной растительности становится меньше определенного числа
от ее общей биомассы. Если биомасса доступной растительности становится меньше определенного числа ![]() , то есть ее леммингам становится энергетически невыгодно и выедание прекращается. Доступная для выедания растительность вычисляется по формуле:
, то есть ее леммингам становится энергетически невыгодно и выедание прекращается. Доступная для выедания растительность вычисляется по формуле:
| (3) |
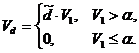
Динамика растительности (V) определяется летом по формуле Ферхюльста, зимой происходит экспоненциальное отмирание, весной прирост и отмирание скомпенсированы.
| (4) |
| (5) |
Здесь ![]() ,
, ![]() – коэффициенты прироста и отмирания биомассы растительности соответственно,
– коэффициенты прироста и отмирания биомассы растительности соответственно, ![]() – емкость экологической ниши.
– емкость экологической ниши.
Отторжение леммингами растительности, с учетом фактора сезонности определяется по формуле:
| (6) |
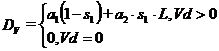
где ![]() ,
, ![]() – коэффициенты отчуждения растительности: первый – зимой и весной и второй – летом.
– коэффициенты отчуждения растительности: первый – зимой и весной и второй – летом.
Прирост биомассы леммингов ![]() вычисляется по формуле:
вычисляется по формуле:
| (7) |
Функция g2 является трофической функцией и отражает снижение ценности кормов при их дефиците.
Коэффициенты ![]() ,
, ![]() - характеризуют сезонные изменения.
- характеризуют сезонные изменения.
Естественная смертность леммингов ![]() зависит от сезона, наличия кормов и определяется по формуле:
зависит от сезона, наличия кормов и определяется по формуле:
1) ![]() , при
, при ![]() ;
;
2) ![]() - при достаточном обеспечении кормами (Vd/L≥q) ;
- при достаточном обеспечении кормами (Vd/L≥q) ;
3) ![]() - при нехватке корма (
- при нехватке корма (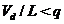 );
);
4) ![]() - при отсутствии корма (
- при отсутствии корма (![]() ), если все съедено весной;
), если все съедено весной;
5) ![]() - при отсутствии корма (
- при отсутствии корма (![]() ), если все съедено уже зимой, , и к концу зимы они остаются только в оптимальном биотопе и численность их там равна
), если все съедено уже зимой, , и к концу зимы они остаются только в оптимальном биотопе и численность их там равна ![]() , а затем численность считается по третьему варианту.
, а затем численность считается по третьему варианту.
Здесь функция ![]() описывает изменение смертности в зависимости от времени года;
описывает изменение смертности в зависимости от времени года; ![]() – критический удельный вес растительности, ниже которого наступает нехватка корма,
– критический удельный вес растительности, ниже которого наступает нехватка корма, ![]() ,
, ![]() ,
, ![]() ,
, ![]() – коэффициенты смертности:
– коэффициенты смертности: ![]() – корма достаточно,
– корма достаточно, ![]() – корма не хватает,
– корма не хватает, ![]() – при отсутствии корма весной.
– при отсутствии корма весной.
Отчуждение леммингов песцами ![]() пропорционально произведению биомасс обоих видов при низкой относительной плотности леммингов (отношение L/F мало) и пропорционально биомассе песцов при высокой плотности (отношение L/F велико):
пропорционально произведению биомасс обоих видов при низкой относительной плотности леммингов (отношение L/F мало) и пропорционально биомассе песцов при высокой плотности (отношение L/F велико):
| (8) |
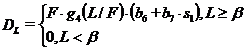
Здесь коэффициенты ![]() ,
, ![]() характеризуют сезонные изменения интенсивности отлова леммингов, функция
характеризуют сезонные изменения интенсивности отлова леммингов, функция 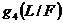 описывает трофическую функцию песцов.
описывает трофическую функцию песцов.
| (9) |
Прирост биомассы песцов в основном обеспечивается леммингами, но кроме них в рационе песцов есть и иные, неспецифические корма, потребление которых считается пропорциональным биомассе песцов.
Прирост биомассы за счет леммингов считается пропорциональным произведению биомассы добытых леммингов на функцию ![]() . Эта функция отражает лимитирующее влияние высокой плотности. Здесь
. Эта функция отражает лимитирующее влияние высокой плотности. Здесь ![]() – коэффициент перехода биомассы леммингов в биомассу песцов, коэффициенты
– коэффициент перехода биомассы леммингов в биомассу песцов, коэффициенты ![]() ,
, ![]() описывают вклад в сезонную изменчивость неспецифического питания.
описывают вклад в сезонную изменчивость неспецифического питания.
Естественная смертность и гибель от охоты считаются пропорциональными биомассе песцов:
| (10) |
| (11) |
где ![]() ,
, ![]() – коэффициенты естественной смертности и интенсивности охоты.
– коэффициенты естественной смертности и интенсивности охоты.
Начальные условия, при которых проводились численные эксперименты:
![]() ,
, ![]() ,
, ![]() 1.
1.
Численные эксперименты проводились при следующих значениях коэффициентов:
Таблица 2. Нормирование экспертных функций.
Функция | Нормирование по Х | Нормирование по Y |
g2 | 100 | 1 |
g3 | - | 16 |
g4 | 22 | 1 |
g5 | 7 | 1 |
Таблица 3. Коэффициенты в модели, при которых проводилось численное исследование
Параметр | Знач. | Описание | Параметр | Знач. | Описание |
Подсистема растительности | |||||
| 0.15 | доля территории, на которой растительность доступна в течение всего года |
| 0.05 | коэффициент выедания растительности зимой, ц/(га × экз. на 100 ловушко-суток × мес.) |
d | 0.6 | надземная часть растительности |
| 0.16 | коэффициент выедания растительности летом, ц/(га × экз. на 100 ловушко-суток × мес.) |
| 2.0 | крит. уровень биомассы, ниже которого выедание становится энергетически не выгодно, ц/га |
| 0.05 | коэффициент отмирания растительности, 1/мес. |
| 30.0 | максимальное значение биомассы растительности, ц/га |
| 1.2 | коэффициент интенсивности прироста растительности, 1/мес. |
подсистема леммингов | |||||
| 500.0 | масштабный коэффициент |
| 0.1 | коэффициент переменной интенсивности выедания леммингов, леммингов на 100 ловушко-суток / (песцов на 100 ловушко-суток × мес.) |
| 0.57 | коэффициент смертности при отсутствии корма (зимой), 1/мес. |
| 0.1 | масштабный коэффициент |
| 0.37 | коэффициент смертности, если корма не хватает, 1/мес. |
| 5.6 | коэффициент перехода биомассы растительности в биомассу леммингов для весны и лета, экз. на 100 ловушко-суток × га/ц |
| 0.05 | коэффициент смертности, если корма хватает, 1/мес. |
| 1.5 | коэффициент перехода биомассы растительности в биомассу леммингов для зимы, экз. на 100 ловушко-суток × га/ц |
| 0.05 | коэффициент постоянной интенсивности выедания леммингов, леммингов на 100 ловушко-суток / (песцов на 100 ловушко-суток × мес.) | q | 0.2 | критический удельный вес растительности, ниже которого наступает нехватка корма |
| 2.5 | уровень оптимального биотопа леммингов, экз. на 100 ловушко-суток | bb | =b2 | Смертность при подрыве кормовой базы |
подсистема песцов | |||||
| 1.00 | коэффициент перехода биомассы леммингов в биомассу песцов, песцов на 100 ловушко-суток / (леммингов на 100 ловушко-суток) |
| 0.055 | коэффициент смертности, 1/мес. |
| 0.025 | коэффициент постоянной интенсивности неспецифического питания, 1/мес. |
| 0.005 | коэффициент интенсивности гибели от охоты, 1/мес. |
| 0.06 | коэффициент сезонной интенсивности неспецифического питания, 1/мес. |
Обратная имитационная задача (построение упрощенной модели).
Для решения «обратной имитационной задачи» – выбора таких постулатов базовой имитационной модели РЛП, которые позволили бы получить разностное уравнение, исходя из результатов вычислительных экспериментов (и в соответствии с эколого-биологическими данными), были проведены следующие упрощения: 1) из рассмотрения была удалена подсистема песцов, т. к. их влияние на динамику леммингов пренебрежимо мало; 2) считалось, что в конце летнего периода биомасса растительности достигает максимального значения ![]() и во все периоды, кроме зимы и весны после пика численности, кормовая база (растительность) не лимитирует динамику численностей леммингов; 3) внутри каждого из сезонов, которые будет характеризовать продолжительностью:
и во все периоды, кроме зимы и весны после пика численности, кормовая база (растительность) не лимитирует динамику численностей леммингов; 3) внутри каждого из сезонов, которые будет характеризовать продолжительностью: 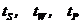 для лета, зимы и весны, соответственно. проводилась кусочная линеаризации нелинейных зависимостей - трофическая функция
для лета, зимы и весны, соответственно. проводилась кусочная линеаризации нелинейных зависимостей - трофическая функция ![]() равна константе
равна константе ![]() - при нехватке корма и
- при нехватке корма и ![]() - корма достаточно; зависимость смертности леммингов от обеспеченности кормами описывалась двумя способами: либо корма достаточно (b5 - минимальная смертность), либо он вообще недоступен (b2 - максимальная смертность). Функция
- корма достаточно; зависимость смертности леммингов от обеспеченности кормами описывалась двумя способами: либо корма достаточно (b5 - минимальная смертность), либо он вообще недоступен (b2 - максимальная смертность). Функция ![]() , отражающая зависимость смертности леммингов от сезона, в каждом из сезонов выбиралась постоянной и равной:
, отражающая зависимость смертности леммингов от сезона, в каждом из сезонов выбиралась постоянной и равной: ![]()
![]() ,
, ![]() – для зимы,
– для зимы, ![]() – для весны,
– для весны, ![]() – для лета.
– для лета.
В соответствии со сделанными предположениями изменения биомассы популяции леммингов в «благоприятный год» - корма достаточно весь год, описывается в каждом из сезонов автономным дифференциальным уравнением первого порядка. В результате, для этого случая, получаем разностное уравнение, связывающее численность леммингов в двух соседних годах,
![]() .
.
Здесь ![]() ,
,![]() – прирост биомассы в благоприятный год;
– прирост биомассы в благоприятный год; 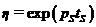 ,
, 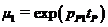 ,
, ![]() – отношение биомасс леммингов в конце и начале сезона, соответственно, летнего, весеннего и зимнего при наличии корма в течение всего сезона;
– отношение биомасс леммингов в конце и начале сезона, соответственно, летнего, весеннего и зимнего при наличии корма в течение всего сезона; 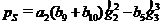
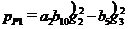 ,
, 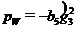 ; параметры
; параметры ![]() и
и ![]() определены в приложении 1.
определены в приложении 1.
При высокой численности леммингов возможно лимитирующее воздействие растительности. В зимний период изменение биомассы растительности описывается по формуле
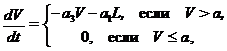
при начальных условиях ![]() , а изменения биомассы популяции леммингов определяется либо по формуле
, а изменения биомассы популяции леммингов определяется либо по формуле
![]()
где, либо, если ![]() то считается, что лемминги остаются только в оптимальном биотопе, т. е. в конце зимы
то считается, что лемминги остаются только в оптимальном биотопе, т. е. в конце зимы ![]() . В свою очередь, в весенний период, в соответствии с предположениями в исходной базовой модели, процессы прироста и отмирания растительности компенсируют друг друга и динамика биомассы растительности определяется воздействиями леммингов
. В свою очередь, в весенний период, в соответствии с предположениями в исходной базовой модели, процессы прироста и отмирания растительности компенсируют друг друга и динамика биомассы растительности определяется воздействиями леммингов
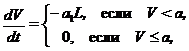
а динамика биомассы популяции леммингов находится по формулам
![]()
Здесь 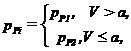
![]() .
.
Используем приведенные выше формулы для получения искомого разностного уравнения. При принятых предположениях оно состоит из трех фрагментов. Первый фрагмент – корма достаточно весь год – происходит линейный рост биомассы (численности) леммингов, третий – корма не хватает уже в зимний период, выживают только особи в оптимальном биотопе – биомасса (численность) постоянна в этом диапазоне (горизонтальный участок – «ступенька») и между ними находится второй фрагмент: переходная зона – корма не хватает в весенний период. Если первый и третий фрагменты имеют простое представление, то при вычислении вида разностного уравнения в переходной зоне получаются громоздкие выражения. Но это никак не оправдано. Как показывают вычислительные эксперименты переходная зона достаточно узкая. Опишем ее отрезком прямой, соединяющим первый и третий фрагменты. Определим условие перехода в третий фрагмент - найдем величину «B» – численность леммингов в конце летнего периода, при которой доступный корм выедается в конце зимы, т. е. в конце зимы биомасса растительности равна ![]() . В силу линейности соответствующего уравнения биомасса растительности аддитивно изменяется от собственных внутрипопуляционных процессов и от выедания леммингами, которое будем считать пропорциональным их средней биомассе. Соответствующее уравнение для определения B имеет вид
. В силу линейности соответствующего уравнения биомасса растительности аддитивно изменяется от собственных внутрипопуляционных процессов и от выедания леммингами, которое будем считать пропорциональным их средней биомассе. Соответствующее уравнение для определения B имеет вид
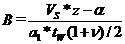 .
.
Здесь 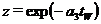 – отношение биомасс растительности в конце и начале зимы при отсутствии воздействия леммингов,
– отношение биомасс растительности в конце и начале зимы при отсутствии воздействия леммингов, ![]() - средняя биомасса леммингов в зимний период.
- средняя биомасса леммингов в зимний период.
Определим теперь ![]() из условия, что корм кончается точно в конце весны.
из условия, что корм кончается точно в конце весны.
Предположим, что в конце лета достигли значения L*. Тогда получаем: численность леммингов в конце зимы равна L* *![]() , а в конце весны равна L* *
, а в конце весны равна L* *![]() и из условия баланса имеет место уравнение для определения L*, которое после учета прироста за лето даст
и из условия баланса имеет место уравнение для определения L*, которое после учета прироста за лето даст![]()
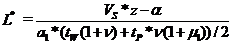 ;
; ![]() = L* *
= L* *![]() .
.
На основании приведенного выше, разностного уравнение, связывающего численность леммингов в двух соседних годах, для нормированной переменной 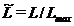 имеет вид:
имеет вид:
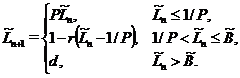
Здесь 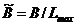 ,
, 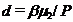 ,
, 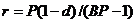 ,
, 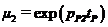 – отношение биомасс леммингов в конце и начале весны при отсутствии корма в течение всего сезона;
– отношение биомасс леммингов в конце и начале весны при отсутствии корма в течение всего сезона; ![]() – биомасса леммингов к концу зимы, если бескормица наступила уже зимой (емкость оптимального биотопа).
– биомасса леммингов к концу зимы, если бескормица наступила уже зимой (емкость оптимального биотопа).
Список необходимых для моделирования параметров.
Количественное описание, как отдельных популяций, так и рассмотрение их в рамках сообществ, требует разнообразной информации. Набор параметров может быть различный. Приводим три набора (с неизбежным некоторым повторением).
Первый список.
Описание популяционных характеристик и факторов среды, на них влияющих.
1. Основные характеристики: функции от плотности, метеоусловий, обеспеченности кормами.
1.1.Продолжительность жизни
1.2.Количество щенков в помете
1.3.Время полового созревания
1.4.Смертность
1.5.Длительность беременности
2. Популяционные характеристики (зависят от плотности, метеоусловий, обеспеченности кормами). Совместный временный ряд.
2.1.Динамика плотности
2.2.Возрастная структура
2.3.Доля размножающихся самок
2.4.Метеоусловия
2.5.Состояние кормовой базы
2.6.Динамика занятости нор
2.7.Стратегия охоты (добывания корма)
3. Временные характеристики популяции
3.1.Время года
3.2.Период появления потомства
3.3.Временный период выкармливания
3.4.«Летний» период, «зимний» период, время «весенней» и «осенней» откочевки
3.5.Критические периоды
4. Пространственные характеристики
4.1.Места норения, их прирученность к биотокам, доля пригодных к норению мест в общей площади территории.
4.2.Размеры индивидуальных участков.
4.3.Оценка подвижности летом, зимой, при откочевках (средняя скорость за сутки, величина обследуемой территории или средний радиус активности).
5.Спектр кормов (с указанием предпочтения в зависимость физиолого-популяционных параметров от состава и качества кормов)
5.1.состав,
5.2.доля кормов – зависимость от сезона и плотности.
6. Биоэнергетические характеристики.
6.1.Характеристики молодых
6.1.1.периоды питания молоком и другими кормами, 6.1.2.обеспечиваемыми родителями
6.1.3.периоды смешанного питания
6.1.4.время самостоятельного питания
6.2. Динамики веса
6.2.1.молодых со времени рождения
6.2. 2.взрослых от сезона, зависимость от погоды и плотности
7. Смертность разных возрастов и разные периоды сезона, зависимость от погоды и плотности
7.1. детально для молодых
8. Энерготраты
8.1.основной обмен
8.2.терморегуляция
8.3.траты энергии и времени на разные виды деятельности
8.3.1.добывание пищи
8.3.2.обустройство нор и территории
8.3.3.перемещения
8.3.4.на охоту территории
8.3.5.прочие виды деятельности
9. Отстрел.
10. Социальная структура, популяция, её зависимость от 3-х факторов и её влияние на физиолого-популяционные характеристики.
11.В каждой точке пространства и в каждый временной такт имеется набор параметров, характеризующих внешнюю среду.
11.1.Температура
11.2.Осадки
11.3.Растительность – биомасса, увлажненность, содержание микроэлементов и т. д. – у каждого вида растительности.
11.4.Характеристика почвенного покрова.
Второй список.
Сбор информации для построения
индивидуально ориентированной модели.
1.Потребности особей
1.1.Движение
1.2.Безопасность
1.3.Кормление
1.4.Продолжение рода
1.5.Выделение отходов метаболизма
1.6.Исследовательская деятельность
1.7.Игры, внутрисемейные контакты
2.Решающие правила
2.1.Начало движения из места покоя (норы)
2.2.Начало поиска места покоя (убежища)
2.3.Критерий контакта
2.4.Исход контакта
2.5.Гибель особи
2.6.Наступления половой зрелости
2.7.Беременности особи
3.Начало движения
(Зависит от статуса особи (возраст, ранг, состояние…), от погоды.)
Может определяться в зависимости от ряда потребностей.
3.1.чувство голода
3.2.потребность в движении
3.3.сексуальном контакте
3.4.Любопытство
3.5.отправление естественных надобностей
3.6.беспокойство за охрану территории (возможна совместная охрана – крики о помощи).
4.Решение по направлению и скорости движения
Это зависит от выбора особью характера движения.
4.1. кормление
4.2. преследование (на своей территории, изгнание «чужаков»).
4.3. Движение (дружеское) к другой особи
4.4. Убегание (от хищников, от преследователей на чужой территории).
4.5. игры
4.6. исследовательская деятельность.
5.Не мотивированное (внешне) движение
5.1.При кормлении скорость стандартная (крейсерская), движение или случайное, или с учетом состояния кормов и расположения других особей.
5.2.Преследование происходит с максимальной скоростью к чужаку.
5.3.Убегание с максимальной скоростью к убежищу (норе).
5.4.Дружеские контакты, игры, исследовательская деятельность – могут быть различные сценарии, в простейшем случае со стандартной скоростью к дружественной особи, а в других, из перечисленных случаев случайное блуждание.
6.Определение возможности контакта
Должна быть задана – зона видимости
7.Исход контакта
Сложна функция от места события и типов особей в контакте.
8.Гибель особи
Зависит от потенциала особи, веса, погодных катаклизмов.
Третий список.
Список основных параметров для моделирования животных (в том числе хищников).
Общие положения. Неформальное введение о физиологии и экологии данного вида.
а). Каждый параметр рассматривается как функция от плотности популяции, климатических (метео) факторов, количества и качественного состава кормов (и иногда от возраста особи и сезона (времени года)).
б). Каждый параметр характеризуется средним, минимальным и максимальным значениями и гистограммой.
1. Эколого-физиологические характеристики
1.1. продолжительность жизни (в виварии, в естественных условиях),
1.2. Количество щенков в помете,
1.3. Длительность беременности,
1.4. Периоды питания: только молоком, молоком и другими кормами, обеспечиваемыми родителями,
1.4.1. Смешанное (самостоятельно + от родителей),
1.4.2. Начало сомастоятельного питания,
1.5. Время распада выводка,
1.6. Время полового созревания,
1.7. Спектр кормов: а. состав, б. доля кормов (обязательно в различные сезоны и зависимость от плотности).
2. Пространственные характеристики.
2.1. места норкения: их приуроченность к биотипам, рельефу местности и т. п. и доля пригодных мест в общей площади,
2.2. Размеры индивидуальных участков,
2.3. Оценка подвижности : летом, зимой, при откочевках (средняя скорость за сутки, величина обследуемой территории или средний радиус активности).
3. Временные характеристики популяции
3.1. время года,
3.2. Период появления потомства,
3.3. Длительность выкармливания,
3.4. «Летний» период, «зимний» период, время «весенней» и «осенней» откочевки
3.5. критические периоды
4.Популяционные характеристики
(необходимо рассмотрение совместное временных рядов, указанных выше ниже факторов и кроме того, зависимость этих факторов от показателей указанных в п 1. общ. положений)
4.1.Динамика плотности популяции,
4.2. Возрастная структура популяции,
4.3. Доля размножающихся самок,
4.4. Смертность,
4.4.1. В различные возрастные периоды (эмбриональная, мертворожденные, у слепых, лактирующих, …., до половой зрелости, в №1,2, и т. д. лет..),
4.4.2. причины смертности: стрессы во время беременности, бескормица (у лактирующих через маломолочность самок), каннибализм родителей, травмы, заболевания.
4.5. Метеоусловия (температура, осадки и др.),
4.6. Состояние кормовой базы (количественное и качественное)
4.7. Динамика занятости нор (или других убежищ),
4.8. Стратегия охоты (добывания корма),
4.9.подвижность особей.
5. Биоэнергетические характеристики
5.1. Динамики веса: молодых (от рождения до половозрелости) и взрослых (как всех др. параметров, функции от сезона, погоды и плотности),
5.2. энерготраты: основной обмен, терморегуляция,
5.3. траты энергии и времени:
5.3.1. добывание пищи,
5.3.2. обустройство нор и территории,
5.3.3. Перемещения,
5.3.4. охрана территории
5.3.5. прочие виды деятельности
5.4.Социальная структура.
Описание структуры-иерархии в различные периоды жизнедеятельности. Её зависимость от факторов пп1) общ. полож. (плотности популяции, климатических факторов, сезона, возрастной структуры и т. д.)
5.5.Механизмы охоты на данный вид
Время промысла, стратегия охоты и их зависимость от различных факторов: плотности погоды и т. д.
, Саранча Д.А.,
Қазақстанның түрлі аймақтарында биогеоценоз математикалық моделдерін құру үшін аралас әдісті қолдану («өсімділік-леммингілер – ақ түлкілер» имитациялық моделі)
Мұнда «өсімділік – леммингілер - ақ түлкілер» иммитациялық моделі қарастырылады. «Кері иммитациялық мәселе» шешіледі. Сонымен бірге жануарларды (соның ішінде жыртқыштарды) моделдеу үшін негізгі параметрлер тізімі берілді.
Boranbayev S. N., Sarancha D. A., Taberkhan R., Trasheev R. V.
Applying of combined methods for initiation of mathematical modeling of biogeocenosis in the different regions of Kazakhstan (imitational models “vegetation – lemming – polar fox”)
The imitational model “vegetation – lemming – polar fox” will be considered. “In verse imitational task” will be solved. The list of main parameters for modeling animals (including predators) is given.
