
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Импульсная активность отдельных нейронов отводилась экстраклеточным способом [15] у животных с выработанным и закрепленным инструментальным поведением. На магнитофон марки "Брюль и Къер-7003" регистрировались: разрядная активность нейрона, звуковой оборонительный сигнал, электрокожное раздражение, отметки нажатия на педаль и взятия пищи из кормушки, актограмма перемещений животного по экспериментальной камере. Для регистрации актограммы на черепе животного укрепляли миниатюрную электролампочку, а на противоположных стенах камеры (возле педали и кормушки) устанавливали фотоэлектрические пластины. По величине суммарной фото-ЭДС на них можно было судить о местоположении животного в тот или иной момент времени. Дополнительно по специальной методике [31] осуществлялась регистрация электромиограммы глубокой части собственно жевательных мышц. После окончания опыта магнитную запись воспроизводили с редукцией скорости в 10 раз на чернилопишущий прибор.
Объектом изучения были нейроны сенсомоторной области коры головного мозга. Координаты места трепанации кости черепа - A3, L3; P3, L3. При выборе зоны отведения учитывалось заключение, что именно на уровне сенсомоторной коры у 60% клеток при формировании мотивации голода наблюдается возникновение характерной пачкообразной активности [17]. Вместе с тем, здесь весьма отчетливо проявляется и другой базовый нейрофизиологический феномен - феномен поведенческой специализации нейронов [1, 3]. Это обстоятельство позволяет считать, что "именно сенсомоторная кора является одним из ответственных пунктов головного мозга, где на основе восходящих мотивационных возбуждений разыгрываются физиологические процессы формирования целенаправленного поведения" [27, стр.9].
Анализ полученных результатов проводился в три этапа. На первом из них с помощью применяемой системы поведенческих отметок на нейронограмме выделяли интервалы времени, соответствующие периодам реализации отдельных инструментальных актов. Для каждого такого периода определяли следующие параметры разрядной деятельности регистрируемого в данный момент нейрона: среднюю частоту его импульсации, структуру фазных активаций (т. е. тип поведенческой специализации клетки) и модальность гистограмм распределения межспайковых интервалов. В последнем случае для нейронов, имеющих низкий уровень разрядной активности построение интервальных гистограмм проводилось путем объединения данных по нескольким (3-5) "смежным" инструментальным актам.
Основной этап обработки полученных данных включал в себя сопоставление показателей средней частоты, специализации и интервальных характеристик одного и того же нейрона в пищедобывательных и оборонительных инструментальных актах. Затем, на основе известных методов регрессионного анализа выясняли, существуют ли направленные изменения перечисленных выше показателей в ходе постепенного снижения уровня мотивации голода по мере насыщения животного. Пищедобывательные и оборонительные действия рассматривались при этом независимо друг от друга.
Достоверность полученных данных оценивали по критерию Стьюдента (уровень значимости р‹0,05). Более подробно методика экспериментов и особенности анализа результатов изложены ранее [8, 9].
Результаты исследования
В общей сложности была исследована разрядная активность 134 нейронов сенсомоторной области коры головного мозга.
Первое, что сразу обращает на себя внимание при рассмотрении полученных данных, - наличие у значительного числа зарегистрированных клеток (49%) закономерно повторяющихся от акта к акту однонаправленных изменений текущей частоты импульсации, приуроченных к строго определенным этапам инструментального действия.
Это явление, хорошо известное как феномен поведенческой специализации нейронов, констатируется в выводах большинства микроэлектродных исследований, проводимых на свободноподвижных животных. При этом отмечается очень высокий уровень стабильности подобных функциональных перестроек [3, 14].
Стабильность феномена поведенческой специализации нейронов отчетливо проявлялась и в рассматриваемом нами случае. Более того, возможность анализировать разрядную активность одной и той же нервной клетки в условиях многократного перехода от пищедобывательного поведения к оборонительному позволяет сделать вывод не только о стабильности, но и об очень быстром и полном восстановлении всей структуры поведенческой специализации нейронов после каждой очередной смены приспособительной направленности инструментального акта. Нам ни разу не удалось наблюдать каких-либо проявлений частичного или хотя бы даже постепенного восстановления паттерна нейронной активности в период возобновления пищедобывательных действий после их прерывания поведением оборонительного характера. Несомненный интерес представляет и тот факт, что фазные изменения частоты импульсации отмечались у изученной нами группы сенсомоторных нейронов именно на стадии реализации приспособительных действий. Ни у одной из этих клеток не удалось зарегистрировать активаций или торможений в ответ на включение звукового оборонительного сигнала или в момент прекращения исходного пищедобывательного поведения.
Вместе с тем, было установлено, что доминирующая мотивация выступает в качестве важного фактора, определяющего структуру специализации нервных клеток. Об этом свидетельствуют результаты сравнительного анализа нейронной активности в сходных по своим двигательным параметрам пищедобывательных и оборонительных инструментальных действиях. Основные результаты такого анализа приведены на рис.3.
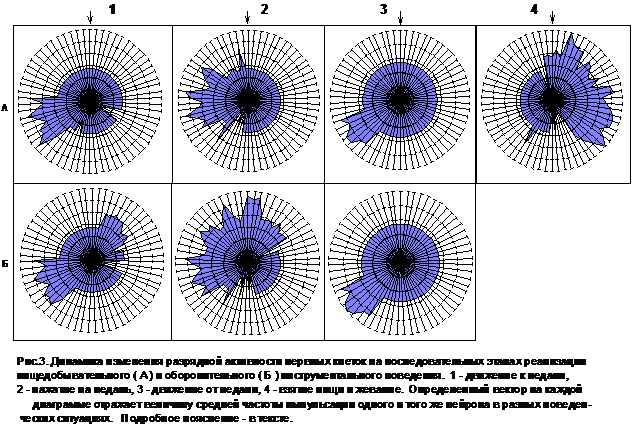
Каждая из семи представленных здесь векторных диаграмм отражает разрядную активность одновременно 66 нейронов, у которых на определенных этапах пищедобывательного (А) и/или оборонительного (Б) инструментального поведения имели место статистически достоверные (р‹0,05) изменения частоты импульсации. Номер вектора на всех диаграммах соответствует индексу одной и той же нервной клетки. Окружность меньшего диаметра условно отражает средний уровень "фоновой" разрядной деятельности нейронов (для разных клеток он, естественно, был различен). Возрастание или, наоборот, снижение частоты генерации потенциалов действия на определенном этапе инструментального акта выражается в сдвигах по определенной оси в сторону от центра (активация) или к центру (торможение). Поскольку амплитуда таких изменений у разных нейронов варьировала в довольно широких пределах, соответствующие величины откладывались по осям в логарифмическом масштабе.
Представленные на рис.3 данные свидетельствуют о том, что нейрональное обеспечение идентичных по способу достижения результата пищедобывательных и оборонительных инструментальных актов было различным. Паттерн импульсации 57% фазно-активных нервных клеток менялся при изменении мотивационной основы поведения.
У 9 нейронов было выявлено статистически достоверное повышение частоты разрядов только в пищедобывательных актах (р‹0,05). В оборонительном поведении импульсная активность у них либо вообще отсутствовала, либо носила регулярный характер. У 3 других клеток наблюдалась фазная активность исключительно в рамках инструментальной оборонительной деятельности. Особо выделялась группа из 42 нейронов, которые избирательно активировались во время развертывания как пищедобывательного, так и оборонительного проведения. Фазные активации 23 из них были связаны с одними и теми же этапами реализации акта нажатия на педаль независимо от его мотивационной основы. Паттерн разрядной активности других 19 нейронов был различен в пищедобывательных и оборонительных инструментальных действиях, но на разных этапах их реализации такого рода различия проявлялись в разной мере. Если во время движения животного к педали и кормушке, как правило, имели место однонаправленные изменения импульсации в пищедобывательном и оборонительном поведении, то в период нажатия животным на педаль такой однонаправленности уже не наблюдалось.
Необходимо особо подчеркнуть, что в рамках каждой из двух сравниваемых форм инструментального поведения амплитуда фазных нейронных активаций или торможений (т. е. величина изменения частоты импульсации по отношению к "фону") существенно варьировала. Нельзя было выделить и двух пищедобывательных или оборонительных актов, нейрональное обеспечение которых по этому критерию было бы полностью идентичным. Причем, проведенный корреляционный анализ показал, что степень выраженности фазных перестроек нейронной активности не зависит от двигательных параметров инструментального действия, в частности от продолжительности различных его этапов, количества и длительности нажатий животным на педаль и т. д. Величина коэффициентов корреляции практически во всех случаях имела значения, которые не достигали критериального уровня р‹0,05.
Кроме того, было установлено, что по мере насыщения животного и, как следствие, снижения уровня пищевой потребности амплитуда активаций (или торможений) нервных клеток также статистически достоверно не меняется. Ни в одном случае нам не удалось выявить корреляционной зависимости между величиной изменения частоты нейронной импульсации на соответствующем этапе поведения и количеством съеденных кроликом порций пищи. Даже в тех случаях, когда длительная регистрация одной и той же нервной клетки сопровождалась плавным возрастанием интенсивности разрядной деятельности (5 нейронов) или, наоборот, ее снижением (4 нейрона), величина фазных изменений частоты импульсации оставалась в пределах статистической погрешности.
Заключение о том, что амплитуда нейрональных активаций не зависит от временных показателей инструментальных действий и не меняется в процессе насыщения подопытных животных имеет принципиально важное значение, поскольку позволяет с единых позиций подходить к анализу всей группы последовательно регистрируемых в опыте нейронов. Т. е. период регистрации каждой клетки можно рассматривать как некоторую “выборку” из одной и той же (генеральной) совокупности аналогичных поведенческих актов [9]. Это, в свою очередь, дает возможность проводить объективную оценку динамики параллельно протекающих в мозге элементарных нейрофизиологических процессов на основе данных, получаемых путем последовательной (выборочной) регистрации каждого из них в отдельности.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
