
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Механизмы мотивационной детерминации
нейронной активности в поведении
С позиций общей теории функциональных систем рассматриваются экспериментальные данные, полученные при проведении сравнительного анализа нейрофизиологических механизмов пищедобывательных и оборонительных инструментальных действий, идентичных по способу достижения результата. Обсуждается круг вопросов, касающихся различных аспектов проблемы мотивационной детерминации нейронной активности в поведении.
Ключевые слова: мотивация, нейроны, функциональная система, целенаправленное поведение, инструментальные формы нейрохимического взаимодействия.
Результаты уже самых первых микроэлектродных исследований, проведенных на бодрствующих не анестезированных животных, позволили установить, что значительная часть нейронов головного мозга млекопитающих обладает спонтанной биоэлектрической активностью [32, 33, 35]. Оказалось, что даже в условиях строго контролируемой сенсорной депривации у клеток зрительной [32], сенсомоторной [2] коры, других отделов центральной нервной системы (ЦНС) [16, 19, 34] все равно наблюдается интенсивная разрядная деятельность.
Рассмотрению этого явления, анализу лежащих в его основе нейрофизиологических механизмов было посвящено множество специальных экспериментальных работ. Но несмотря на большой объем выполненных в данном направлении исследований, причины возникновения спонтанной нейронной активности и сегодня еще остаются до конца не ясными.
Попытки получить ответ на этот вопрос на основе представлений о процессах "внутреннего" возбуждения ЦНС человека и животных дали довольно противоречивые результаты. С одной стороны, действительно были установлены факты, свидетельствующие в пользу гипотезы о существовании особых подкорковых структур, нейроны которых при изменении определенных показателей гомеостаза способны оказывать генерализованное возбуждающее влияние на клетки вышележащих отделов головного мозга [36]. На первый взгляд, удалось даже идентифицировать конкретные паттерны нейронной активности, специфичные в отношении процессов формирования пищевой [24, 30], питьевой [21], ряда других метаболических потребностей организма [6, 22].
Однако результаты последующих экспериментальных работ, выполненных на базе более совершенных методов регистрации и анализа нейронной активности, не подтвердили основные выводы предыдущих исследований. Так, по данным ряда авторов никакого значительного возрастания средней частоты разрядов клеток подкорковых структур мозга при возникновении внутренней потребности на самом деле не происходит [18, с.170]. А это значит, что нейроны указанных отделов ЦНС не могут в такие периоды времени выполнять функцию пейсмекеров, т. е. оказывать прямое возбуждающее влияние на какие-либо другие клеточные единицы.
Не менее серьезные вопросы возникают и при рассмотрении данных, указывающих на специфичность нейрональных паттернов мотивационного возбуждения. Дело в том, что во всех исследованиях, посвященных решению этой проблемы, соответствующие заключения основывались на результатах анализа интервальных гистограмм, построенных в системе координат с неравномерной шкалой времени. Ширина канала составляла 10 мс (для интервалов от 0 до 100 мс) и 100 мс (для интервалов в области 100-1000 мс). Утверждалось, что возникновение пищевой [7, 13, 29], питьевой [21] или оборонительной [23, 28] мотивации сопровождается массовым переходом нервных клеток от регулярной формы разрядной активности к характерному пачечно-групповому типу импульсации. В качестве критерия доминирования “пачкообразной” ритмики рассматривалось наличие двух экстремумов ("бимодальность") соответствующих интервальных гистограмм.
Нетрудно понять, что использование подобного методического приема на самом деле полностью исключает возможность объективного решения поставленной задачи. В этом можно убедиться на следующем простом примере.
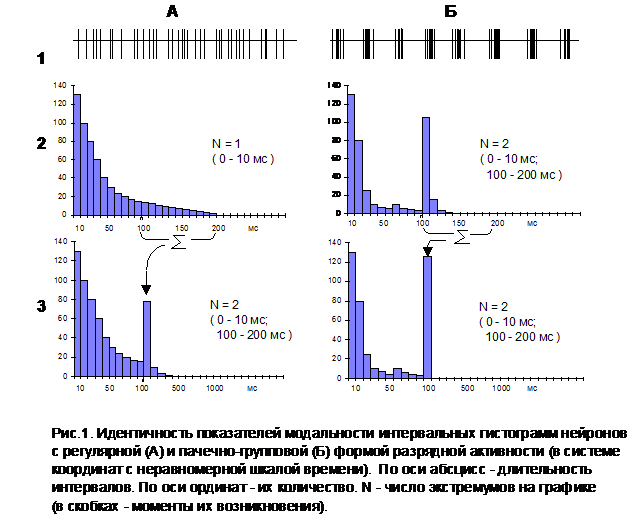
На рис.1А представлена нейронограмма, распределение межспайковых интервалов которой аппроксимируется классической мономодальной кривой (А2). Ниже - та же самая гистограмма, но построенная уже в системе координат с неравномерной шкалой времени (А3). Сравнивая эти графики легко заметить, что до момента изменения масштаба оси абсцисс они в точности повторяют друг друга. Однако, начиная со 100 мсек такое соответствие нарушается. Каждому очередному каналу гистограммы А3 начинает соответствовать суммарное "содержимое" десяти каналов исходного распределения (А2). В результате образуется второй (фантомный) пик в области 100-200 мс, и нейрон с четко выраженной регулярной формой активности начинает ошибочно идентифицироваться как клетка пачечно-группового типа.
Проделав аналогичное преобразование для нейронов, действительно обладающих "пачкообразной" конфигурацией биоэлектрических разрядов (рис.1Б), и сопоставив полученные графики, приходится признать, что в системах координат с неравномерной шкалой времени переход от регулярного типа импульсации (рис.1.А3) к пачечно-групповой (рис.1.Б3) не поддается объективизации по критерию модальности интервальных гистограмм. На первый взгляд, не понятно, каким образом здесь вообще удается получать какие-либо мономодальные распределения.
Ответ дает следующий простой пример.
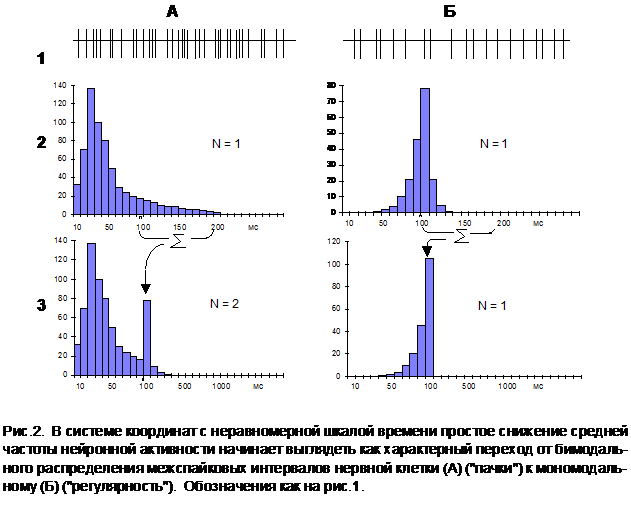
На рис.2 представлены интервальные гистограммы нейрона, обладающего регулярной формой разрядной активности, но имеющего различную интенсивность импульсации в разные периоды времени. Хорошо видно, что если средняя величина межспайковых интервалов находится в пределах 100-200 мс, соответствующее распределение сохраняет свой мономодальный вид и после преобразования (Б2—›Б3). Но при этом оказывается, что в системе координат с неравномерной шкалой времени в зависимости от средней частоты импульсной активности один и тот же нейрон может быть идентифицирован как клетка и "пачечно-группового" (рис.3А), и регулярного (рис.3Б) типа.
Подводя определенный итог вышесказанному, приходится с сожалением констатировать, что результаты проведенных к настоящему времени исследований не позволяют сделать вывод о существовании каких-либо специфических нейрональных кодов, отражающих на клеточном уровне процесс формирования мотивационного возбуждения. На основе применяемой авторами процедуры первичной обработки данных в принципе невозможно провести объективное разделение всей совокупности регистрируемых нервных клеток на нейроны с регулярной и пачечно-групповой формой импульсной активности. Этот вывод, естественно, распространяется и на утверждение о наличии особого, мотивационно-детерминированного распределения по отделам головного мозга нейронов с характерной пачкообразной ритмикой (90% - латеральный гипоталамус, 60% - сенсомоторная кора, 45% - зрительная область коры и т. п.) [17]. На самом деле, даже если число таких клеток будет равно нулю, наличие феномена бимодальности интервальных гистограмм все равно вполне может отмечаться в тех же самых соотношениях (рис.1А).
Кроме того, ни в одной из проведенных в данном направлении многочисленных работ нельзя найти каких-либо математических выкладок (или, хотя бы, доверительных интервалов на гистограммах), которые на основе результатов аналитической статистики давали бы надежду другим исследователям полагать, что повторив применяемую авторами последовательность экспериментальных процедур, они также с достаточно высокой вероятностью будут наблюдать именно описываемый ряд закономерностей.
Если с учетом вышеизложенного попытаться сформулировать перечень требований, соблюдение которых является необходимым для проведения корректных исследований в этом направлении, следует выделить следующий ряд основных моментов:
1. Применяемые экспериментальные модели должны предоставлять возможность неоднократного повторения в ходе регистрации активности каждой исследуемой нервной клетки ситуации контролируемого изменения уровня или биологической модальности доминирующей мотивации при наличии строгих физиологических и поведенческих критериев смены соответствующих функциональных состояний организма.
2. Нейрофизиологические закономерности, наблюдаемые в рамках любого конкретного интервала времени, следует рассматривать только в качестве выборочных данных из соответствующей генеральной совокупности. Это является необходимым условием проведения полноценного статистического анализа, включающего в себя определение достоверности наблюдаемых нейрональных феноменов.
3. Постановка экспериментов должна осуществляться только на свободно подвижных животных, поскольку в настоящее время установлено большое число фактов, свидетельствующих о кардинальном изменении функциональных свойств нервных клеток в условиях иммобилизации. В последнем случае по сути дела приходится говорить об особом разделе нейрофизиологии, в котором констатируются закономерности, имеющие мало общего с реальными процессами, протекающими в нервной системе интактного, свободноподвижного животного.
4. Применяемые экспериментальные модели должны давать возможность рассмотрения получаемых данных на основе простых однофакторных схем анализа, поскольку в естественных условиях контролировать весь комплекс переменных, влияющих на исследуемый нейрофизиологический процесс, как правило, не удается. Переход же ко все более и более жестким аналитическим схемам проведения опыта неизбежно приводит к искажению реальной картины изучаемых явлений.
Цель настоящей работы - рассмотреть возможные пути решения данного круга вопросов на основе современных методов экспериментального исследования.
Методика
Перечисленному выше ряду требований в полной мере удовлетворяет разработанная в нашей лаборатории экспериментальная модель, в рамках которой подопытные животные (кролики) во время регистрации у них активности отдельных нервных клеток выполняли нажатие передними лапами на площадку специальной педали для получения пищи и устранения по звуковому сигналу болевого электрокожного раздражения (ЭКР) [8]. Наличие высокого уровня оборонительной мотивации в последнем случае подтверждается результатами электрокардиографического анализа [12].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
