
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Как показывают результаты такого анализа, нейроны, обладающие определенными типами поведенческой специализации (рис.3), действительно являются компонентами одной и той же функциональной системы, в рамках которой активность всех ее составляющих характеризуется очень высоким уровнем временной синхронизации. Узловыми моментами проявления такого рода синхронизации, как правило, выступают определенные события организменного уровня, связанные с достижением животным этапных или конечных результатов. А это значит, что по основным своим признакам указанная группа нейронов полностью соответствует критерию системоспецифичности [4, 5].
Совершенно иная картина складывается, если попытаться оценить динамику тех же самых системных процессов на основе анализа интервальных характеристик нейронной активности. Как показывает проведенный анализ, лишь у 14% сенсомоторных нейронов синхронно возникают характерные пачечно-групповые конфигурации биоэлектрических разрядов, специфичные в отношении процесса формирования пищедобывательного поведения [7, 13, 24 и др.]. Для большинства же клеток (84%) вообще не удается установить наличие каких бы то ни было общих интервальных показателей, свидетельствующих о согласованном характере их разрядной деятельности. По сути дела здесь приходится говорить о соотношении: один нейрон - один паттерн.
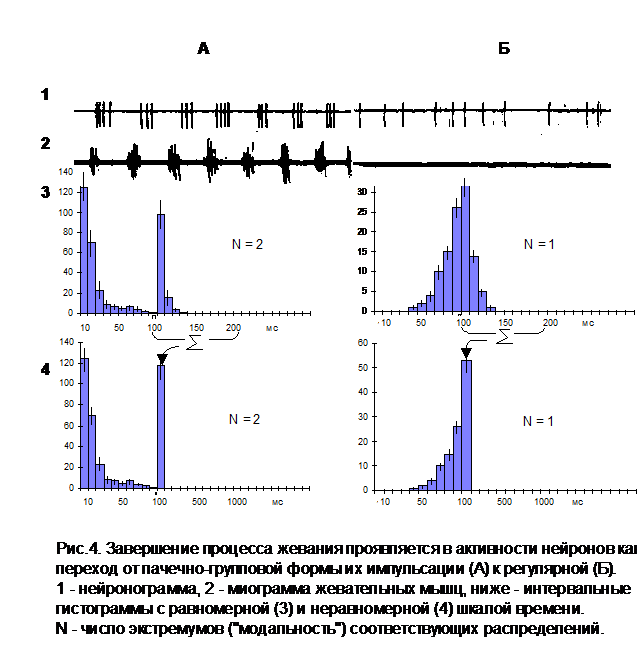
Но даже для первой группы клеток (14% нейронов) синхронное возникновение пачечно-групповой формы импульсной активности, как выяснилось, не связано с процессом формирования мотивации голода, а обусловлено особенностями их поведенческой специализации. Оказалось, что все они относятся к числу “нейронов жевания” (рис.4). В пользу этого заключения свидетельствует строгая корреляция их разрядной деятельности с определенной фазой жевательных движений. Кроме того, по мере насыщения животного у данной группы клеток ни разу не отмечалось постепенного снижения степени выраженности феномена "пачкообразной" ритмики при жевании. Между тем, отдельные нейроны нам удавалось регистрировать непрерывно в течение нескольких часов во время неоднократно воспроизводимых ситуаций удовлетворения пищевой потребности и ее повторного возникновения.
Если в свете этих данных вновь обратиться к примеру, представленному на рис.4, нетрудно понять, что в рамках простых экспериментальных моделей, когда наличие мотивации голода и факт ее удовлетворения могут быть объективно определены только по критерию поедания животным пищи или отказу от нее, “нейроны жевания” неизбежно будут ошибочно идентифицироваться как нейроны соответствующего мотивационного возбуждения. А поскольку число таких клеток значительно больше (свыше 90% [1]) в центральных структурах мозга, очень легко сделать и другой ошибочный вывод - вывод о существовании градуального распределения нейронов пищевой мотивации по различным отделам ЦНС [17].
Обсуждение результатов
Результаты проведенных нами экспериментальных исследований определенно свидетельствуют о том, что в период реализации целенаправленного инструментального поведения доминирующая мотивация не играет роль императивного фактора, способного оказывать прямое влияние на процесс формирования разрядной активности отдельных нервных клеток. Это заключение касается как показателей средней частоты, так и конфигурации (паттерна) нейронных разрядов. Основным проявлением мотивационной специфичности работы нервных клеток является определенная структура их поведенческой специализации, которая существенным образом меняется при смене характера приспособительной направленности инструментальных действий. Отмечаемый же в публикациях ряда авторов феномен массового возникновения “пачкообразной” (мотивационно-детерминированной) формы нейронной импульсации является артефактом первичной обработки данных и обусловлен неправомерным применением системы координат с неравномерной шкалой времени при построении соответствующих интервальных гистограмм (рис.1).
В наиболее последовательной форме полученные данные могут быть интерпретированы с позиций основных положений общей теории функциональных систем [4]. Согласно развиваемым здесь представлениям, разрядная активность нейронов головного мозга в естественных условиях не формируется как реакция на синаптические воздействия, поступающие со стороны рецепторов или клеток других структур ЦНС (в том числе и “мотивациогенных” подкорковых центров). Она является следствием особенностей функционального статуса определенной части нейронов, наличия у них собственных пейсмекерных механизмов, связанных с развертыванием базовых внутриклеточных функций. Соответственно, объединение такого рода фоновоактивных нейронов в единую интеграцию реализуется как процесс синхронного устранения "избыточного числа степеней свободы" [5] (рис.5А).
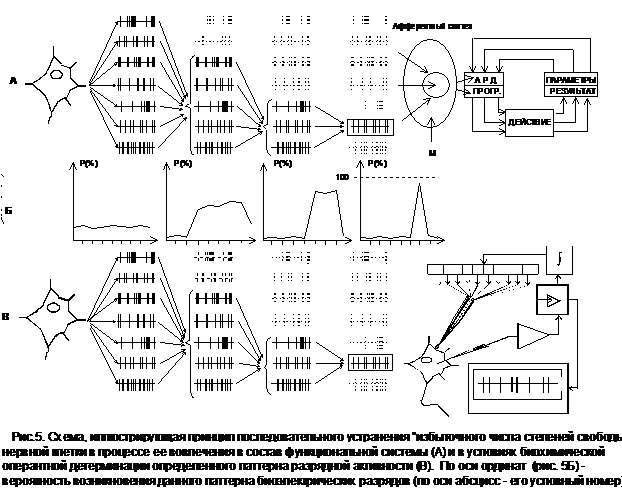
С этих позиций нейрофизиологическую основу поведения человека и животных составляют процессы формирования совершенно иных по набору своих компонентов центрально-периферических интеграций, нежели те, которые можно объективизировать по критерию эффективного синаптического межклеточного взаимодействия. Последнее рассматривается лишь как один из многих механизмов вовлечения отдельного компонента в состав соответствующей функциональной системы. Ведущий же принцип построения каждой такой интеграции определяется как принцип взаимосодействия всех ее составляющих на получение полезного для организма приспособительного результата [4, С.41]. Вследствие этого именно результат становится императивным фактором, определяющим процесс устранения избыточного числа степеней свободы нервных клеток (рис.5А). Любые же другие системные детерминанты, включая пусковой стимул или доминирующую мотивацию, подобным свойством не обладают.
Принципиально важное значение при этом приобретает вопрос о роли синаптических механизмов в процессах формирования нейронной активности. То, что решающая роль здесь принадлежит именно нейромедиаторым функциям не вызывает сомнений. Но вот конкретная схема, в рамках которой происходит их реализация, - во многом еще остается не определенной.
Острота этой проблемы вполне понятна. Как уже отмечалось, активность нейронов, входящих в состав любой функциональной системы, характеризуется исключительно высоким уровнем стабильности [1, 14]. И по своей структуре, и по степени выраженности закономерно повторяющиеся от акта к акту фазные нейронные активации сохраняются не только при значительных изменениях различных параметров двигательной активности животного, но даже в случае кардинальной реорганизации некоторых основных условий достижения приспособительного результата [2, 19]. Обращает на себя внимание и отсутствие направленной динамики изменения фазных перестроек нейронной импульсации в ходе развертывания большого числа идентичных целенаправленных действий.
Попытка достижения такого уровня стабильности на основе обычных ("стимуляционных") подходов к моделированию нейромедиаторных функций неизбежно заканчивается неудачей. Одна из причин - быстрое развитие процесса привыкания (габитуации) нейрона к любым внешним воздействиям [25]. Такого рода габитуационная форма динамики вызванной нейронной активности отражает базовое функциональное свойство любой нервной клетки и наблюдается даже в случае прямой микроионофоретической аппликации возбуждающих нейромедиаторов [20] или при деполяризации клеточной мембраны [26]. Причем, чем ближе параметры нейромедиаторного воздействия к естественным показателям (строгая локальность зоны действия вещества, кратковременность), тем менее устойчивой становится ответная нейронная реакция [10]. Как же в таком случае возникает феномен поведенческой специализации нервных клеток?
Контуры определенного ответа на этот вопрос стали вырисовываться после того, как было установлено, что наряду с классическими ("хемореактивными") формами нейромедиаторного взаимодействия в ЦНС человека и животных возможна принципиально иная схема проявления биологически активного характера веществ медиаторной природы (рис.5В). Оказалось, что они могут выступать не только в качестве инициаторов развития хемореактивных процессов, но и как факторы подкрепления эндогенно - или экзогенно-детерминированных перестроек импульсации фоновоактивных нейронов [11]. Причем, динамика подобных функциональных изменений на клеточном уровне, а также условия их возникновения качественным образом отличаются от аналогичных параметров хемореактивных процессов. Но особого внимания, конечно же, заслуживает высокий уровень устойчивости достигаемых таким путем перестроек нейронной активности.
С этих позиций исключительно важное значение для проведения дальнейших исследований в данном направлении приобретает вопрос о базовом нейрофизиологическом феномене, составляющем основу процесса обучения. Как известно, современные нейрофизиологические концепции строго ориентированы в своих заключениях на вывод об изменении эффективности синаптической передачи. На наш взгляд, схема детерминации нейронной активности по (рис.5А) может быть реализована только на основе совершенно иного принципа модификации элементарных нейрофизиологических функций при обучении. Весьма вероятно, что решающая роль здесь принадлежит именно процессам реорганизации естественных форм инструментального нейрохимического взаимодействия. С нашей точки зрения, детальный анализ этого феномена, выяснение лежащих в его основе молекулярно-биологических механизмов имеет принципиально важное значение для разработки новых базовых нейрофизиологических моделей, учитывающих способность нервных клеток объединяться в устойчивые по составу группы (функциональные системы), в рамках которых строго согласованный характер активности отдельных элементов достигается на основе процесса положительного биохимического подкрепления индивидуальных паттернов разрядной деятельности.
В Ы В О Д Ы
Основной формой проявления мотивационной специфичности работы нейронов головного мозга в поведении является определенная структура их системной специализации, которая существенным образом меняется при изменении характера приспособительной направленности инструментальных действий.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
